 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 7jij | ||||||
|---|---|---|---|---|---|---|---|
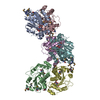
| Title | ATP-bound AMP-activated protein kinase | ||||||
 Components Components |
| ||||||
 Keywords Keywords |  SIGNALING PROTEIN / AMPK / activation / ATP-binding SIGNALING PROTEIN / AMPK / activation / ATP-binding | ||||||
| Function / homology |  Function and homology information Function and homology informationnegative regulation of glucosylceramide biosynthetic process / positive regulation of mitochondrial transcription / [hydroxymethylglutaryl-CoA reductase (NADPH)] kinase / [hydroxymethylglutaryl-CoA reductase (NADPH)] kinase activity / : / regulation of stress granule assembly / histone H2BS36 kinase activity / AMPK inhibits chREBP transcriptional activation activity / regulation of peptidyl-serine phosphorylation / cold acclimation ...negative regulation of glucosylceramide biosynthetic process / positive regulation of mitochondrial transcription / [hydroxymethylglutaryl-CoA reductase (NADPH)] kinase / [hydroxymethylglutaryl-CoA reductase (NADPH)] kinase activity / : / regulation of stress granule assembly / histone H2BS36 kinase activity / AMPK inhibits chREBP transcriptional activation activity / regulation of peptidyl-serine phosphorylation / cold acclimation / positive regulation of peptidyl-lysine acetylation / lipid droplet disassembly / Lipophagy / regulation of bile acid secretion / positive regulation of skeletal muscle tissue development / CAMKK-AMPK signaling cascade / import into nucleus / cAMP-dependent protein kinase regulator activity / regulation of vesicle-mediated transport / positive regulation of cholesterol biosynthetic process / cellular response to organonitrogen compound / nucleotide-activated protein kinase complex / Energy dependent regulation of mTOR by LKB1-AMPK / Carnitine metabolism / negative regulation of hepatocyte apoptotic process / tau-protein kinase / bile acid and bile salt transport / protein kinase regulator activity / cellular response to ethanol / protein localization to lipid droplet / negative regulation of TOR signaling / bile acid signaling pathway / Activation of PPARGC1A (PGC-1alpha) by phosphorylation / response to caffeine / motor behavior / regulation of glycolytic process / positive regulation of protein targeting to mitochondrion / cAMP-dependent protein kinase activity / lipid biosynthetic process / negative regulation of tubulin deacetylation / AMP-activated protein kinase activity / Macroautophagy / positive regulation of protein localization / tau-protein kinase activity / AMP binding / cholesterol biosynthetic process / fatty acid oxidation / cellular response to nutrient levels / carbohydrate transmembrane transporter activity / positive regulation of protein kinase activity / fatty acid homeostasis / negative regulation of lipid catabolic process / cellular response to glucose starvation / positive regulation of autophagy / energy homeostasis / Activation of AMPK downstream of NMDARs / regulation of microtubule cytoskeleton organization / response to UV / negative regulation of TORC1 signaling / positive regulation of adipose tissue development / cellular response to calcium ion / negative regulation of insulin receptor signaling pathway / positive regulation of glycolytic process / response to activity / Translocation of SLC2A4 (GLUT4) to the plasma membrane / TP53 Regulates Metabolic Genes / response to gamma radiation / cellular response to glucose stimulus / tau protein binding / regulation of circadian rhythm / ADP binding / fatty acid biosynthetic process / autophagy / Wnt signaling pathway / cellular response to hydrogen peroxide / neuron cellular homeostasis / response to estrogen / cellular response to prostaglandin E stimulus / glucose metabolic process / rhythmic process / cellular response to xenobiotic stimulus / glucose homeostasis / positive regulation of cold-induced thermogenesis / cellular response to oxidative stress / cellular response to hypoxia / spermatogenesis / Regulation of TP53 Activity through Phosphorylation / periplasmic space / non-specific serine/threonine protein kinase / response to hypoxia / protein kinase activity / nuclear speck / apical plasma membrane / axon / protein phosphorylation / negative regulation of gene expression / protein serine kinase activity / protein serine/threonine kinase activity / neuronal cell body / dendriteSimilarity search - Function | ||||||
| Biological species |  Escherichia coli (E. coli) Escherichia coli (E. coli) Homo sapiens (human) Homo sapiens (human) | ||||||
| Method | X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 5.5 Å | ||||||
 Authors Authors | Yan, Y. / Zhou, X.E. / Powell, K. / Xu, T. / Brunzelle, J.S. / Xu, H.X. / Melcher, K. | ||||||
| Funding support |  United States, 1items United States, 1items
| ||||||
 Citation Citation | Journal: Science / Year: 2021 Title: Structure of an AMPK complex in an inactive, ATP-bound state. Authors: Yan Yan / Somnath Mukherjee / Kaleeckal G Harikumar / Timothy S Strutzenberg / X Edward Zhou / Kelly Suino-Powell / Ting-Hai Xu / Ryan D Sheldon / Jared Lamp / Joseph S Brunzelle / Katarzyna ...Authors: Yan Yan / Somnath Mukherjee / Kaleeckal G Harikumar / Timothy S Strutzenberg / X Edward Zhou / Kelly Suino-Powell / Ting-Hai Xu / Ryan D Sheldon / Jared Lamp / Joseph S Brunzelle / Katarzyna Radziwon / Abigail Ellis / Scott J Novick / Irving E Vega / Russell G Jones / Laurence J Miller / H Eric Xu / Patrick R Griffin / Anthony A Kossiakoff / Karsten Melcher /  Abstract: Adenosine monophosphate (AMP)-activated protein kinase (AMPK) regulates metabolism in response to the cellular energy states. Under energy stress, AMP stabilizes the active AMPK conformation, in ...Adenosine monophosphate (AMP)-activated protein kinase (AMPK) regulates metabolism in response to the cellular energy states. Under energy stress, AMP stabilizes the active AMPK conformation, in which the kinase activation loop (AL) is protected from protein phosphatases, thus keeping the AL in its active, phosphorylated state. At low AMP:ATP (adenosine triphosphate) ratios, ATP inhibits AMPK by increasing AL dynamics and accessibility. We developed conformation-specific antibodies to trap ATP-bound AMPK in a fully inactive, dynamic state and determined its structure at 3.5-angstrom resolution using cryo-electron microscopy. A 180° rotation and 100-angstrom displacement of the kinase domain fully exposes the AL. On the basis of the structure and supporting biophysical data, we propose a multistep mechanism explaining how adenine nucleotides and pharmacological agonists modulate AMPK activity by altering AL phosphorylation and accessibility. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 7jij.cif.gz | 506.4 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb7jij.ent.gz | 415.3 KB | Display | PDB format |
| PDBx/mmJSON format | 7jij.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/ji/7jijftp://data.pdbj.org/pub/pdb/validation_reports/ji/7jij | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  7jhgC  7jhhC  7m74C  4rerS S: Starting model for refinement C: citing same article ( |
|---|---|
| Similar structure data |




-Links
 PDBj
PDBj













- Assembly
Assembly
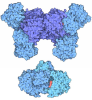
| Deposited unit | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||
| Unit cell |
|
-Components
-Protein , 2 types, 2 molecules MA
| #1: Protein | Mass: 40827.125 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Escherichia coli (E. coli) / Gene: malE, GRW33_05015 / Production host:   Spodoptera frugiperda (fall armyworm) / References: UniProt: A0A6D0N546 Spodoptera frugiperda (fall armyworm) / References: UniProt: A0A6D0N546 |
|---|---|
| #2: Protein | Mass: 56084.375 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: PRKAA1, AMPK1 / Production host: Spodoptera frugiperda (fall armyworm)References: UniProt: Q13131, non-specific serine/threonine protein kinase, [acetyl-CoA carboxylase] kinase, [hydroxymethylglutaryl-CoA reductase (NADPH)] kinase, tau-protein kinase |
-5'-AMP-activated protein kinase subunit ... , 2 types, 2 molecules BG
| #3: Protein | Mass: 22384.650 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: PRKAB2 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: O43741 |
|---|---|
| #4: Protein | Mass: 34893.453 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: PRKAG1 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: P54619 |
-Sugars , 1 types, 1 molecules
| #5: Polysaccharide | alpha-D-glucopyranose-(1-4)-alpha-D-glucopyranose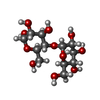  , Oligosaccharide / Class: Nutrient / Mass: 342.297 Da / Num. of mol.: 1 , Oligosaccharide / Class: Nutrient / Mass: 342.297 Da / Num. of mol.: 1Source method: isolated from a genetically manipulated source Details: oligosaccharide / References: alpha-maltose |
|---|
-Non-polymers , 3 types, 3 molecules 

| #6: Chemical | ChemComp-ATP / Adenosine triphosphate Mass: 507.181 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Comment: ATP, energy-carrying molecule*YM Mass: 507.181 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H16N5O13P3 / Comment: ATP, energy-carrying molecule*YM |
|---|---|
| #7: Chemical | ChemComp-AMP / Adenosine monophosphate Mass: 347.221 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H14N5O7P / Comment: AMP*YM Mass: 347.221 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H14N5O7P / Comment: AMP*YM |
| #8: Chemical | ChemComp-ADP / Adenosine diphosphate Mass: 427.201 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Comment: ADP, energy-carrying molecule*YM Mass: 427.201 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Comment: ADP, energy-carrying molecule*YM |
-Details
| Has ligand of interest | Y |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 5 Å3/Da / Density % sol: 75.39 % |
|---|---|
| Crystal grow | Temperature: 293 K / Method: vapor diffusion Details: 0.2 M tri-lithium citrate,10% w/v PEG3350, pH7.5,0.01 M barium chloride |
-Data collection
| Diffraction | Mean temperature: 100 K / Serial crystal experiment: N |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: APS / Beamline: 21-ID-D / Wavelength: 1.07823 Å |
| Detector | Type: DECTRIS EIGER2 X 9M / Detector: PIXEL / Date: Jul 5, 2017 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 1.07823 Å / Relative weight: 1 |
| Reflection | Resolution: 5.5→49 Å / Num. obs: 10563 / % possible obs: 99.5 % / Redundancy: 9.7 % / CC1/2: 0.979 / Rmerge(I) obs: 0.298 / Net I/σ(I): 4.7 |
| Reflection shell | Resolution: 5.5→6.15 Å / Rmerge(I) obs: 3.1 / Num. unique obs: 2894 / CC1/2: 0.323 |
- Processing
Processing
| Software |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: 4RER Resolution: 5.5→24.901 Å / SU ML: 1.02 / Cross valid method: THROUGHOUT / σ(F): 1.34 / Phase error: 43.94 / Stereochemistry target values: ML
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.8 Å / VDW probe radii: 1.1 Å / Solvent model: FLAT BULK SOLVENT MODEL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso max: 971.36 Å2 / Biso mean: 429.4287 Å2 / Biso min: 166.08 Å2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: final / Resolution: 5.5→24.901 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell | Refine-ID: X-RAY DIFFRACTION / Rfactor Rfree error: 0 / Total num. of bins used: 4
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS params. | Method: refined / Refine-ID: X-RAY DIFFRACTION
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS group |
|