 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 3d4g | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | ZP-N domain of mammalian sperm receptor ZP3 (crystal form II) | |||||||||
 Components Components | Maltose-binding periplasmic protein, LINKER, Zona pellucida protein 3 | |||||||||
 Keywords Keywords |  CELL ADHESION / fertilization / oocyte / egg coat / zona pellucida / vitelline envelope / ZP domain / egg-sperm interaction / species-specific gamete recognition / speciation / biodiversity / infertility / extracellular matrix / immunoglobulin-like fold / glycoprotein / receptor / secreted / transmembrane CELL ADHESION / fertilization / oocyte / egg coat / zona pellucida / vitelline envelope / ZP domain / egg-sperm interaction / species-specific gamete recognition / speciation / biodiversity / infertility / extracellular matrix / immunoglobulin-like fold / glycoprotein / receptor / secreted / transmembrane | |||||||||
| Function / homology |  Function and homology information Function and homology informationpositive regulation of type IV hypersensitivity / positive regulation of acrosomal vesicle exocytosis / positive regulation of antral ovarian follicle growth / Interaction With Cumulus Cells And The Zona Pellucida / egg coat formation / positive regulation of humoral immune response / structural constituent of egg coat / egg coat / acrosin binding / negative regulation of binding of sperm to zona pellucida ...positive regulation of type IV hypersensitivity / positive regulation of acrosomal vesicle exocytosis / positive regulation of antral ovarian follicle growth / Interaction With Cumulus Cells And The Zona Pellucida / egg coat formation / positive regulation of humoral immune response / structural constituent of egg coat / egg coat / acrosin binding / negative regulation of binding of sperm to zona pellucida / positive regulation of acrosome reaction / positive regulation of ovarian follicle development / binding of sperm to zona pellucida / humoral immune response mediated by circulating immunoglobulin / oocyte development / blastocyst formation / regulation of signaling receptor activity / positive regulation of leukocyte migration / detection of maltose stimulus / maltose binding / maltose transport complex / maltose transport / maltodextrin transmembrane transport / carbohydrate transmembrane transporter activity / ATP-binding cassette (ABC) transporter complex, substrate-binding subunit-containing / carbohydrate transport / positive regulation of interleukin-4 production / positive regulation of T cell proliferation / ATP-binding cassette (ABC) transporter complex / cell chemotaxis / extracellular matrix / positive regulation of inflammatory response / positive regulation of type II interferon production / outer membrane-bounded periplasmic space / carbohydrate binding / collagen-containing extracellular matrix / periplasmic space / receptor ligand activity / negative regulation of DNA-templated transcription / DNA damage response / positive regulation of DNA-templated transcription / extracellular space / membrane / identical protein binding / plasma membraneSimilarity search - Function | |||||||||
| Biological species |  Escherichia coli (E. coli) Escherichia coli (E. coli) Mus musculus (house mouse) Mus musculus (house mouse) | |||||||||
| Method | X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.3 Å | |||||||||
 Authors Authors | Jovine, L. / Monne, M. | |||||||||
 Citation Citation | Journal: Nature / Year: 2008 Title: Crystal structure of the ZP-N domain of ZP3 reveals the core fold of animal egg coats Authors: Monne, M. / Han, L. / Schwend, T. / Burendahl, S. / Jovine, L. #1: Journal: CELL(CAMBRIDGE,MASS.) / Year: 1980 Title: Mammalian sperm-egg interaction: identification of a glycoprotein in mouse egg zonae pellucidae possessing receptor activity for sperm Authors: Bleil, J.D. / Wassarman, P.M. #2: Journal: Febs Lett. / Year: 1992 Title: A large domain common to sperm receptors (Zp2 and Zp3) and TGF-beta type III receptor Authors: Bork, P. / Sander, C. #3: Journal: Nat Cell Biol / Year: 2002 Title: The ZP domain is a conserved module for polymerization of extracellular proteins. Authors: Luca Jovine / Huayu Qi / Zev Williams / Eveline Litscher / Paul M Wassarman /  Abstract: Many eukaryotic extracellular proteins share a sequence of unknown function, called the zona pellucida (ZP) domain. Among these proteins are the mammalian sperm receptors ZP2 and ZP3, non-mammalian ...Many eukaryotic extracellular proteins share a sequence of unknown function, called the zona pellucida (ZP) domain. Among these proteins are the mammalian sperm receptors ZP2 and ZP3, non-mammalian egg coat proteins, Tamm-Horsfall protein (THP), glycoprotein-2 (GP-2), alpha- and beta-tectorins, transforming growth factor (TGF)-beta receptor III and endoglin, DMBT-1 (deleted in malignant brain tumour-1), NompA (no-mechanoreceptor-potential-A), Dumpy and cuticlin-1 (refs 1,2). Here, we report that the ZP domain of ZP2, ZP3 and THP is responsible for polymerization of these proteins into filaments of similar supramolecular structure. Most ZP domain proteins are synthesized as precursors with carboxy-terminal transmembrane domains or glycosyl phosphatidylinositol (GPI) anchors. Our results demonstrate that the C-terminal transmembrane domain and short cytoplasmic tail of ZP2 and ZP3 are not required for secretion, but are essential for assembly. Finally, we suggest a molecular basis for dominant human hearing disorders caused by point mutations within the ZP domain of alpha-tectorin. #4: Journal: Proc Natl Acad Sci U S A / Year: 2004 Title: A duplicated motif controls assembly of zona pellucida domain proteins. Authors: Luca Jovine / Huayu Qi / Zev Williams / Eveline S Litscher / Paul M Wassarman / Abstract: Many secreted eukaryotic glycoproteins that play fundamental roles in development, hearing, immunity, and cancer polymerize into filaments and extracellular matrices through zona pellucida (ZP) ...Many secreted eukaryotic glycoproteins that play fundamental roles in development, hearing, immunity, and cancer polymerize into filaments and extracellular matrices through zona pellucida (ZP) domains. ZP domain proteins are synthesized as precursors containing C-terminal propeptides that are cleaved at conserved sites. However, the consequences of this processing and the mechanism by which nascent proteins assemble are unclear. By microinjection of mutated DNA constructs into growing oocytes and mammalian cell transfection, we have identified a conserved duplicated motif [EHP (external hydrophobic patch)/IHP (internal hydrophobic patch)] regulating the assembly of mouse ZP proteins. Whereas the transmembrane domain (TMD) of ZP3 can be functionally replaced by an unrelated TMD, mutations in either EHP or IHP do not hinder secretion of full-length ZP3 but completely abolish its assembly. Because mutants truncated before the TMD are not processed, we conclude that the conserved TMD of mammalian ZP proteins does not engage them in specific interactions but is essential for C-terminal processing. Cleavage of ZP precursors results in loss of the EHP, thereby activating secreted polypeptides to assemble by using the IHP within the ZP domain. Taken together, these findings suggest a general mechanism for assembly of ZP domain proteins. #5: Journal: Annu Rev Biochem / Year: 2005 Title: Zona pellucida domain proteins. Authors: Luca Jovine / Costel C Darie / Eveline S Litscher / Paul M Wassarman / Abstract: Many eukaryotic proteins share a sequence designated as the zona pellucida (ZP) domain. This structural element, present in extracellular proteins from a wide variety of organisms, from nematodes to ...Many eukaryotic proteins share a sequence designated as the zona pellucida (ZP) domain. This structural element, present in extracellular proteins from a wide variety of organisms, from nematodes to mammals, consists of approximately 260 amino acids with eight conserved cysteine (Cys) residues and is located close to the C terminus of the polypeptide. ZP domain proteins are often glycosylated, modular structures consisting of multiple types of domains. Predictions can be made about some of the structural features of the ZP domain and ZP domain proteins. The functions of ZP domain proteins vary tremendously, from serving as structural components of egg coats, appendicularian mucous houses, and nematode dauer larvae, to serving as mechanotransducers in flies and receptors in mammals and nonmammals. Generally, ZP domain proteins are present in filaments and/or matrices, which is consistent with the role of the domain in protein polymerization. A general mechanism for assembly of ZP domain proteins has been presented. It is likely that the ZP domain plays a common role despite its presence in proteins of widely diverse functions. #6: Journal: Bmc Biochem. / Year: 2006 Title: The PLAC1-homology region of the ZP domain is sufficient for protein polymerisation Authors: Jovine, L. / Janssen, W.G. / Litscher, E.S. / Wassarman, P.M. | |||||||||
| History |
| |||||||||
| Remark 650 | HELIX DETERMINATION METHOD: AUTHOR DETERMINED | |||||||||
| Remark 700 | SHEET DETERMINATION METHOD: AUTHOR DETERMINED | |||||||||
| Remark 40 | MOLPROBITY STRUCTURE VALIDATION PROGRAMS: MOLPROBITY (KING, REDUCE, AND PROBE) AUTHORS: I.W.DAVIS,V. ...MOLPROBITY STRUCTURE VALIDATION PROGRAMS: MOLPROBITY (KING, REDUCE, AND PROBE) AUTHORS: I.W.DAVIS,V.B.CHEN, R.M.IMMORMINO, J.J.HEADD,W.B.ARENDALL,J.M.WORD AUTHORS: I.W.DAVIS,A.LEAVER-FAY,V.B.CHEN,J.N.BLOCK, G.J.KAPRAL,X.WANG,L.W.MURRAY,W.B.ARENDALL, J.SNOEYINK,J.S.RICHARDSON,D.C.RICHARDSON REFERENCE: MOLPROBITY: ALL-ATOM CONTACTS AND STRUCTURE VALIDATION FOR PROTEINS AND NUCLEIC ACIDS NUCLEIC ACIDS RESEARCH. 2007;35:W375-83. |
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 3d4g.cif.gz | 1.5 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb3d4g.ent.gz | 1.2 MB | Display | PDB format |
| PDBx/mmJSON format | 3d4g.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/d4/3d4gftp://data.pdbj.org/pub/pdb/validation_reports/d4/3d4g | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  3d4cSC  3ef7C  5osqC S: Starting model for refinement C: citing same article ( |
|---|---|
| Similar structure data |










-Links
 PDBj
PDBj




- Assembly
Assembly
| Deposited unit | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||
| 2 | 
| ||||||||
| 3 | 
| ||||||||
| 4 | 
| ||||||||
| 5 | 
| ||||||||
| 6 | 
| ||||||||
| 7 | 
| ||||||||
| 8 | 
| ||||||||
| Unit cell |
|
-Components
| #1: Protein | Mass: 52948.945 Da / Num. of mol.: 8 Fragment: ZP3 ZP-N domain, UNP residues 27-393, UNP residues 42-143 Mutation: I3T, E360A, K363A, D364A, R368N Source method: isolated from a genetically manipulated source Details: This protein is a chimera. Residues 2-368 are from E. COLI maltose binding protein (MBP), correspond to residues 27-393 of swiss-prot database entry P0AEX9 and contain mutations I3T, E360A, ...Details: This protein is a chimera. Residues 2-368 are from E. COLI maltose binding protein (MBP), correspond to residues 27-393 of swiss-prot database entry P0AEX9 and contain mutations I3T, E360A, K363A, D364A, R368N (corresponding to I28T, E385A, K388A, D389A AND R393N in P0AEX9). Residues 372-473 are from mouse ZP3 protein and correspond to residues 42-143 of swiss-prot database entry P10761. Source: (gene. exp.) Escherichia coli (strain K12) (bacteria), (gene. exp.) Mus musculus (house mouse)Cellular location: Extracellular matrix / Gene: Zp3, Zp-3, Zpc / Plasmid: PLJMBP4C, PLJDIS1 / Production host: Escherichia coli (E. coli) / Strain (production host): Origami B (DE3) / References: UniProt: P0AEX9, UniProt: P10761#2: Polysaccharide | alpha-D-glucopyranose-(1-4)-alpha-D-glucopyranose / alpha-maltose 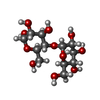  , Oligosaccharide / Class: Nutrient / Mass: 342.297 Da / Num. of mol.: 8 , Oligosaccharide / Class: Nutrient / Mass: 342.297 Da / Num. of mol.: 8Source method: isolated from a genetically manipulated source Details: oligosaccharide / References: alpha-maltose #3: Chemical | ChemComp-CA /   Mass: 40.078 Da / Num. of mol.: 24 / Source method: obtained synthetically / Formula: Ca Mass: 40.078 Da / Num. of mol.: 24 / Source method: obtained synthetically / Formula: Ca#4: Water | ChemComp-HOH / | Water Mass: 18.015 Da / Num. of mol.: 1934 / Source method: isolated from a natural source / Formula: H2O Mass: 18.015 Da / Num. of mol.: 1934 / Source method: isolated from a natural source / Formula: H2O |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 2.76 Å3/Da / Density % sol: 55.48 % |
|---|---|
| Crystal grow | Temperature: 293 K / Method: vapor diffusion, hanging drop / pH: 7 Details: Sample: 15mg/ml protein in 0.05M sodium chloride, 0.01M Tris-HCL, PH7.2, 0.001M maltose. Reservoir: 10% PEG6000, 0.2M calcium chloride, 0.1M Tris-HCL, PH7.0. Sample to reservoir ratio in ...Details: Sample: 15mg/ml protein in 0.05M sodium chloride, 0.01M Tris-HCL, PH7.2, 0.001M maltose. Reservoir: 10% PEG6000, 0.2M calcium chloride, 0.1M Tris-HCL, PH7.0. Sample to reservoir ratio in drop: 1:1, pH7.0, VAPOR DIFFUSION, HANGING DROP, temperature 293.0K |
-Data collection
| Diffraction | Mean temperature: 100 K |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: ESRF  / Beamline: BM14 / Wavelength: 0.9535 / Wavelength: 0.9535 Å / Beamline: BM14 / Wavelength: 0.9535 / Wavelength: 0.9535 Å |
| Detector | Type: MAR CCD 165 mm / Detector: CCD / Date: Dec 7, 2006 |
| Radiation | Monochromator: SI(111) MONOCHROMATOR / Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 0.9535 Å / Relative weight: 1 |
| Reflection | Resolution: 2.3→500 Å / Num. all: 202456 / Num. obs: 193240 / % possible obs: 95.4 % / Observed criterion σ(F): 0 / Observed criterion σ(I): -3 / Redundancy: 2 % / Biso Wilson estimate: 48.357 Å2 / Rsym value: 0.05 / Net I/σ(I): 11 |
| Reflection shell | Resolution: 2.3→2.35 Å / Redundancy: 2 % / Mean I/σ(I) obs: 1.6 / Num. unique all: 12935 / Rsym value: 0.529 / % possible all: 95.7 |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: PDB ENTRY 3D4C Resolution: 2.3→50 Å / Cor.coef. Fo:Fc: 0.959 / Cor.coef. Fo:Fc free: 0.944 / SU B: 15.285 / SU ML: 0.196 / Cross valid method: THROUGHOUT / σ(F): 0 / ESU R: 0.324 / ESU R Free: 0.215 / Stereochemistry target values: MAXIMUM LIKELIHOOD / Details: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Ion probe radii: 0.8 Å / Shrinkage radii: 0.8 Å / VDW probe radii: 1.2 Å / Solvent model: BABINET MODEL WITH MASK | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 50.67 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 2.3→50 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell | Resolution: 2.3→2.36 Å / Total num. of bins used: 20
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS params. | Method: refined / Refine-ID: X-RAY DIFFRACTION
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS group |
|