 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 9it0 | ||||||
|---|---|---|---|---|---|---|---|
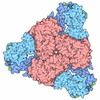
| Title | Liganded-state E.coli PatZ | ||||||
 Components Components | Protein acetyltransferase | ||||||
 Keywords Keywords | TRANSFERASE / Acetyltransferase | ||||||
| Function / homology |  Function and homology information Function and homology informationacyltransferase activity, transferring groups other than amino-acyl groups / ATP binding / metal ion binding Similarity search - Function | ||||||
| Biological species |  | ||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 1.99 Å | ||||||
 Authors Authors | Park, J.B. / Roh, S.H. | ||||||
| Funding support |  Korea, Republic Of, 1items Korea, Republic Of, 1items
| ||||||
 Citation Citation | Journal: Proc Natl Acad Sci U S A / Year: 2025 Title: Structural basis of the catalytic and allosteric mechanism of bacterial acetyltransferase PatZ. Authors: Jun Bae Park / Gwanwoo Lee / Yu-Yeon Han / Dongwook Kim / Kyoo Heo / Jeesoo Kim / Juhee Park / Hyosuk Yun / Chul Won Lee / Hyun-Soo Cho / Jong-Seo Kim / Martin Steinegger / Yeong-Jae Seok / Soung-Hun Roh / Abstract: GCN5-related -acetyltransferases (GNATs) are essential for regulating bacterial metabolism by acetylating specific target proteins. Despite their importance in bacterial physiology, the mechanisms ...GCN5-related -acetyltransferases (GNATs) are essential for regulating bacterial metabolism by acetylating specific target proteins. Despite their importance in bacterial physiology, the mechanisms behind their enzymatic and regulatory functions remain poorly understood. In this study, we investigated the structures of protein acetyltransferase Z (PatZ), a Type I GNAT, and examined its ligand interactions, catalytic mechanism, and allosteric regulation. PatZ functions as a homotetramer, with each subunit comprising a catalytic and a regulatory domain. Our results demonstrate that the regulatory domain is vital for acetyltransferase activity, as it triggers cooperative conformational changes in the catalytic domain and directly aids in the formation of substrate-binding pockets. Additionally, a protein structure-based evolutionary analysis of bacterial GNAT types revealed a distinct regulatory domain pattern across phyla, highlighting its crucial role in responding to cellular energy levels. #1: Journal: mBio / Year: 2018 Title: Identification of Novel Protein Lysine Acetyltransferases in Escherichia coli. Authors: David G Christensen / Jesse G Meyer / Jackson T Baumgartner / Alexandria K D'Souza / William C Nelson / Samuel H Payne / Misty L Kuhn / Birgit Schilling / Alan J Wolfe /  Abstract: Posttranslational modifications, such as ε-lysine acetylation, regulate protein function. ε-lysine acetylation can occur either nonenzymatically or enzymatically. The nonenzymatic mechanism uses ...Posttranslational modifications, such as ε-lysine acetylation, regulate protein function. ε-lysine acetylation can occur either nonenzymatically or enzymatically. The nonenzymatic mechanism uses acetyl phosphate (AcP) or acetyl coenzyme A (AcCoA) as acetyl donor to modify an ε-lysine residue of a protein. The enzymatic mechanism uses ε-lysine acetyltransferases (KATs) to specifically transfer an acetyl group from AcCoA to ε-lysine residues on proteins. To date, only one KAT (YfiQ, also known as Pka and PatZ) has been identified in Here, we demonstrate the existence of 4 additional KATs: RimI, YiaC, YjaB, and PhnO. In a genetic background devoid of all known acetylation mechanisms (most notably AcP and YfiQ) and one deacetylase (CobB), overexpression of these putative KATs elicited unique patterns of protein acetylation. We mutated key active site residues and found that most of them eliminated enzymatic acetylation activity. We used mass spectrometry to identify and quantify the specificity of YfiQ and the four novel KATs. Surprisingly, our analysis revealed a high degree of substrate specificity. The overlap between KAT-dependent and AcP-dependent acetylation was extremely limited, supporting the hypothesis that these two acetylation mechanisms play distinct roles in the posttranslational modification of bacterial proteins. We further showed that these novel KATs are conserved across broad swaths of bacterial phylogeny. Finally, we determined that one of the novel KATs (YiaC) and the known KAT (YfiQ) can negatively regulate bacterial migration. Together, these results emphasize distinct and specific nonenzymatic and enzymatic protein acetylation mechanisms present in bacteria.ε-Lysine acetylation is one of the most abundant and important posttranslational modifications across all domains of life. One of the best-studied effects of acetylation occurs in eukaryotes, where acetylation of histone tails activates gene transcription. Although bacteria do not have true histones, ε-lysine acetylation is prevalent; however, the role of these modifications is mostly unknown. We constructed an strain that lacked both known acetylation mechanisms to identify four new ε-lysine acetyltransferases (RimI, YiaC, YjaB, and PhnO). We used mass spectrometry to determine the substrate specificity of these acetyltransferases. Structural analysis of selected substrate proteins revealed site-specific preferences for enzymatic acetylation that had little overlap with the preferences of the previously reported acetyl-phosphate nonenzymatic acetylation mechanism. Finally, YiaC and YfiQ appear to regulate flagellum-based motility, a phenotype critical for pathogenesis of many organisms. These acetyltransferases are highly conserved and reveal deeper and more complex roles for bacterial posttranslational modification. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 9it0.cif.gz | 1.2 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb9it0.ent.gz | 1 MB | Display | PDB format |
| PDBx/mmJSON format | 9it0.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/it/9it0ftp://data.pdbj.org/pub/pdb/validation_reports/it/9it0 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  60853MC  9isbC  9isqC C: citing same article ( M: map data used to model this data |
|---|---|
| Similar structure data |









-Links
 PDBj
PDBj


- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
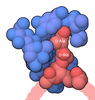
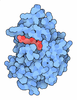
| #1: Protein | Mass: 98099.375 Da / Num. of mol.: 4 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Gene: pat, yfiQ, ACU57_10170, BGM66_000644, BJI68_02105, C0P57_000002, CG831_000342, CIG67_15390, CTR35_002231, CV83915_03530, DTL43_02095, EIZ93_15170, FOI11_000020, FOI11_20030, FWK02_03775, G3V95_ ...Gene: pat, yfiQ, ACU57_10170, BGM66_000644, BJI68_02105, C0P57_000002, CG831_000342, CIG67_15390, CTR35_002231, CV83915_03530, DTL43_02095, EIZ93_15170, FOI11_000020, FOI11_20030, FWK02_03775, G3V95_05750, G4A38_06870, G4A47_19960, GNW61_15560, GOP25_16215, GQM21_00025, GRW05_03440, HMV95_11440, J0541_000369, JNP96_20710, QDW62_06300, SAMEA3752557_00912 Production host: #2: Chemical | ChemComp-ACO /   Mass: 809.571 Da / Num. of mol.: 8 / Source method: obtained synthetically / Formula: C23H38N7O17P3S / Feature type: SUBJECT OF INVESTIGATION Mass: 809.571 Da / Num. of mol.: 8 / Source method: obtained synthetically / Formula: C23H38N7O17P3S / Feature type: SUBJECT OF INVESTIGATION#3: Chemical | ChemComp-PO4 /   Mass: 94.971 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: PO4 / Feature type: SUBJECT OF INVESTIGATION Mass: 94.971 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: PO4 / Feature type: SUBJECT OF INVESTIGATION#4: Water | ChemComp-HOH / |  Mass: 18.015 Da / Num. of mol.: 507 / Source method: isolated from a natural source / Formula: H2O Mass: 18.015 Da / Num. of mol.: 507 / Source method: isolated from a natural source / Formula: H2OHas ligand of interest | Y | Has protein modification | N | |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: Acetyltransferase / Type: COMPLEX / Entity ID: #1 / Source: RECOMBINANT |
|---|---|
| Source (natural) | Organism: |
| Source (recombinant) | Organism: |
| Buffer solution | pH: 7.4 |
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: TFS KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: DARK FIELD / Nominal defocus max: 8000 nm / Nominal defocus min: 1500 nm |
| Image recording | Electron dose: 50 e/Å2 / Film or detector model: FEI FALCON IV (4k x 4k) |
- Processing
Processing
| CTF correction | Type: NONE |
|---|---|
| 3D reconstruction | Resolution: 1.99 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 5121944 / Symmetry type: POINT |
