 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
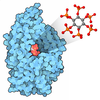
| Entry | Database: PDB / ID: 9fpz | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Human NatA-MAP2 80S ribosome complex | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | TRANSLATION / co-translational processing / ribosome associated factor (RAF) / methionine aminopeptidase 2 (MAP2) / N-terminal methionine excision (NME) / N-acetyl-transferase A (NatA) / N-termional acetylation (NTA) / UBA domain / a-solenoid / protein-protein and protein-RNA interactions | |||||||||
| Function / homology |  Function and homology information Function and homology informationnegative regulation of maintenance of mitotic sister chromatid cohesion, centromeric / protein-N-terminal-glutamate acetyltransferase activity / N-terminal amino-acid Nalpha-acetyltransferase NatA / N-terminal protein amino acid acetylation / NatA complex / protein N-terminal-serine acetyltransferase activity / protein-N-terminal-alanine acetyltransferase activity / protein-N-terminal amino-acid acetyltransferase activity / internal protein amino acid acetylation / N-acetyltransferase activity ...negative regulation of maintenance of mitotic sister chromatid cohesion, centromeric / protein-N-terminal-glutamate acetyltransferase activity / N-terminal amino-acid Nalpha-acetyltransferase NatA / N-terminal protein amino acid acetylation / NatA complex / protein N-terminal-serine acetyltransferase activity / protein-N-terminal-alanine acetyltransferase activity / protein-N-terminal amino-acid acetyltransferase activity / internal protein amino acid acetylation / N-acetyltransferase activity / methionyl aminopeptidase / initiator methionyl aminopeptidase activity / metalloexopeptidase activity / acetyltransferase activator activity / eukaryotic 80S initiation complex / axial mesoderm development / regulation of translation involved in cellular response to UV / 90S preribosome assembly / positive regulation of intrinsic apoptotic signaling pathway in response to DNA damage by p53 class mediator / positive regulation of DNA damage response, signal transduction by p53 class mediator / TORC2 complex binding / regulation of translational initiation / middle ear morphogenesis / protein acetylation / metalloaminopeptidase activity / Peptide chain elongation / Selenocysteine synthesis / Formation of a pool of free 40S subunits / Eukaryotic Translation Termination / Response of EIF2AK4 (GCN2) to amino acid deficiency / SRP-dependent cotranslational protein targeting to membrane / Viral mRNA Translation / Nonsense Mediated Decay (NMD) independent of the Exon Junction Complex (EJC) / GTP hydrolysis and joining of the 60S ribosomal subunit / chromosome organization / L13a-mediated translational silencing of Ceruloplasmin expression / aminopeptidase activity / Major pathway of rRNA processing in the nucleolus and cytosol / protein-RNA complex assembly / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / rough endoplasmic reticulum / cytosolic ribosome / ossification / maturation of LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / positive regulation of translation / ribosomal large subunit biogenesis / skeletal system development / DNA damage response, signal transduction by p53 class mediator / sensory perception of sound / cellular response to gamma radiation / Regulation of expression of SLITs and ROBOs / mRNA 5'-UTR binding / protein processing / cytoplasmic ribonucleoprotein granule / rRNA processing / cellular response to UV / Inactivation, recovery and regulation of the phototransduction cascade / regulation of translation / ribosome binding / cell body / angiogenesis / transcription regulator complex / cytosolic large ribosomal subunit / cytoplasmic translation / cell differentiation / postsynaptic density / protein stabilization / rRNA binding / nuclear body / structural constituent of ribosome / cadherin binding / translation / ribonucleoprotein complex / focal adhesion / mRNA binding / intracellular membrane-bounded organelle / synapse / dendrite / negative regulation of apoptotic process / regulation of DNA-templated transcription / positive regulation of DNA-templated transcription / nucleolus / endoplasmic reticulum / DNA binding / RNA binding / extracellular exosome / nucleoplasm / metal ion binding / nucleus / membrane / plasma membrane / cytosol / cytoplasm Similarity search - Function | |||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | |||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 2.69 Å | |||||||||
 Authors Authors | Klein, M.A. / Wild, K. / Sinning, I. | |||||||||
| Funding support |  Germany, 1items Germany, 1items
| |||||||||
 Citation Citation | Journal: Nat Commun / Year: 2024 Title: Multi-protein assemblies orchestrate co-translational enzymatic processing on the human ribosome. Authors: Marius Klein / Klemens Wild / Irmgard Sinning / Abstract: Nascent chains undergo co-translational enzymatic processing as soon as their N-terminus becomes accessible at the ribosomal polypeptide tunnel exit (PTE). In eukaryotes, N-terminal methionine ...Nascent chains undergo co-translational enzymatic processing as soon as their N-terminus becomes accessible at the ribosomal polypeptide tunnel exit (PTE). In eukaryotes, N-terminal methionine excision (NME) by Methionine Aminopeptidases (MAP1 and MAP2), and N-terminal acetylation (NTA) by N-Acetyl-Transferase A (NatA), is the most common combination of subsequent modifications carried out on the 80S ribosome. How these enzymatic processes are coordinated in the context of a rapidly translating ribosome has remained elusive. Here, we report two cryo-EM structures of multi-enzyme complexes assembled on vacant human 80S ribosomes, indicating two routes for NME-NTA. Both assemblies form on the 80S independent of nascent chain substrates. Irrespective of the route, NatA occupies a non-intrusive 'distal' binding site on the ribosome which does not interfere with MAP1 or MAP2 binding nor with most other ribosome-associated factors (RAFs). NatA can partake in a coordinated, dynamic assembly with MAP1 through the hydra-like chaperoning function of the abundant Nascent Polypeptide-Associated Complex (NAC). In contrast to MAP1, MAP2 completely covers the PTE and is thus incompatible with NAC and MAP1 recruitment. Together, our data provide the structural framework for the coordinated orchestration of NME and NTA in protein biogenesis. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 9fpz.cif.gz | 811 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb9fpz.ent.gz | Display | PDB format | |
| PDBx/mmJSON format | 9fpz.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Summary document | 9fpz_validation.pdf.gz | 1.9 MB | Display | wwPDB validaton report |
|---|---|---|---|---|
| Full document | 9fpz_full_validation.pdf.gz | 1.9 MB | Display | |
| Data in XML | 9fpz_validation.xml.gz | 110.6 KB | Display | |
| Data in CIF | 9fpz_validation.cif.gz | 167.2 KB | Display | |
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/fp/9fpzftp://data.pdbj.org/pub/pdb/validation_reports/fp/9fpz | HTTPS FTP |
-Related structure data
| Related structure data |  50641MC  9fq0C M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |

-Links
 PDBj
PDBj






































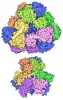





- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-N-alpha-acetyltransferase ... , 2 types, 2 molecules 2B
| #1: Protein | Mass: 20003.795 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: NAA10, ARD1, ARD1A, TE2 / Production host:   Spodoptera frugiperda (fall armyworm) Spodoptera frugiperda (fall armyworm)References: UniProt: P41227, N-terminal amino-acid Nalpha-acetyltransferase NatA |
|---|---|
| #4: Protein | Mass: 98658.648 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: NAA15, GA19, NARG1, NATH, TBDN100 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: Q9BXJ9 |
-RNA chain , 2 types, 2 molecules 81
| #2: RNA chain | Mass: 18646.127 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) |
|---|---|
| #5: RNA chain | Mass: 1640222.125 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) |
-Protein , 1 types, 1 molecules A
| #3: Protein | Mass: 53413.945 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: METAP2, MNPEP, P67EIF2 / Production host: Spodoptera frugiperda (fall armyworm) / References: UniProt: P50579, methionyl aminopeptidase |
|---|
-60S ribosomal protein ... , 6 types, 6 molecules LCLkLhLXLRLr
| #6: Protein | Mass: 47804.621 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P36578 |
|---|---|
| #8: Protein | Mass: 8238.948 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P63173 |
| #10: Protein | Mass: 14462.429 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P42766 |
| #11: Protein | Mass: 17740.193 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P62750 |
| #12: Protein | Mass: 23535.281 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P84098 |
| #13: Protein | Mass: 15784.622 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P46779 |
-Large ribosomal subunit protein ... , 2 types, 2 molecules LELY
| #7: Protein | Mass: 32810.176 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: Q02878 |
|---|---|
| #9: Protein | Mass: 17174.184 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P61254 |
-Non-polymers , 2 types, 3 molecules 
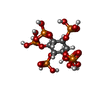
| #14: Chemical |  Mass: 58.933 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Co Mass: 58.933 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Co#15: Chemical | ChemComp-IHP / |  Mass: 660.035 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C6H18O24P6 Mass: 660.035 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C6H18O24P6 |
|---|
-Details
| Has ligand of interest | N |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: human NatA and MAP2 bound to the PTE of the 80S ribosome Type: RIBOSOME / Entity ID: #1-#13 / Source: NATURAL |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Buffer solution | pH: 7.5 |
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: TFS KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal defocus max: 2300 nm / Nominal defocus min: 1300 nm |
| Image recording | Electron dose: 41.28 e/Å2 / Film or detector model: GATAN K3 (6k x 4k) |
- Processing
Processing
| EM software | Name: PHENIX / Version: 1.21_5207: / Category: model refinement | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: NONE | ||||||||||||||||||||||||
| 3D reconstruction | Resolution: 2.69 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 25404 / Symmetry type: POINT | ||||||||||||||||||||||||
| Refine LS restraints |
|
