 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 9fjs | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
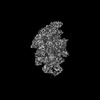
| タイトル | Cryo-EM structure of Mycobacterium tuberculosis sigma-B RNA polymerase bound to -10 promoter element ssDNA oligo - sigma-B undocked conformation | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | TRANSCRIPTION / RNA polymerase / sigma factor / SigB / stress response / -10 promoter element / promoter recognition / tuberculosis | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報RNA polymerase core enzyme binding / Antimicrobial action and antimicrobial resistance in Mtb / sigma factor activity / bacterial-type RNA polymerase core enzyme binding / cytosolic DNA-directed RNA polymerase complex / peptidoglycan-based cell wall / DNA-directed RNA polymerase complex / DNA-templated transcription initiation / ribonucleoside binding / DNA-directed RNA polymerase ...RNA polymerase core enzyme binding / Antimicrobial action and antimicrobial resistance in Mtb / sigma factor activity / bacterial-type RNA polymerase core enzyme binding / cytosolic DNA-directed RNA polymerase complex / peptidoglycan-based cell wall / DNA-directed RNA polymerase complex / DNA-templated transcription initiation / ribonucleoside binding / DNA-directed RNA polymerase / DNA-directed RNA polymerase activity / response to heat / response to hypoxia / protein dimerization activity / response to xenobiotic stimulus / response to antibiotic / DNA-templated transcription / positive regulation of DNA-templated transcription / magnesium ion binding / DNA binding / zinc ion binding / plasma membrane / cytoplasm / cytosol 類似検索 - 分子機能 | ||||||||||||
| 生物種 |  Mycobacterium tuberculosis H37Rv (結核菌) Mycobacterium tuberculosis H37Rv (結核菌) | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.48 Å | ||||||||||||
 データ登録者 データ登録者 | Brodolin, K. / Blaise, M. | ||||||||||||
| 資金援助 |  フランス, 3件 フランス, 3件
| ||||||||||||
 引用 引用 | ジャーナル: Nucleic Acids Res / 年: 2025 タイトル: Single-stranded DNA drives σ subunit loading onto mycobacterial RNA polymerase to unlock initiation-competent conformations. 著者: Rishi Kishore Vishwakarma / Nils Marechal / Zakia Morichaud / Mickaël Blaise / Emmanuel Margeat / Konstantin Brodolin / 要旨: Initiation of transcription requires the formation of the "open" promoter complex (RPo). For this, the σ subunit of bacterial RNA polymerase (RNAP) binds to the nontemplate strand of the -10 element ...Initiation of transcription requires the formation of the "open" promoter complex (RPo). For this, the σ subunit of bacterial RNA polymerase (RNAP) binds to the nontemplate strand of the -10 element sequence of promoters and nucleates DNA unwinding. This is accompanied by a cascade of conformational changes on RNAP, the exact mechanics of which remains elusive. Here, using single-molecule Förster resonance energy transfer and cryo-electron microscopy, we explored the conformational landscape of RNAP from the human pathogen Mycobacterium tuberculosis upon binding to a single-stranded DNA (ssDNA) fragment that includes the -10 element sequence (-10 ssDNA). We found that like the transcription activator RNAP-binding protein A, -10 ssDNA induced σ subunit loading onto the DNA/RNA channels of RNAP. This triggered RNAP clamp closure and unswiveling that are required for RPo formation and RNA synthesis initiation. Our results reveal a mechanism of ssDNA-guided RNAP maturation and identify the σ subunit as a regulator of RNAP conformational dynamics. #1: ジャーナル: Acta Crystallogr D Struct Biol / 年: 2018 タイトル: Real-space refinement in PHENIX for cryo-EM and crystallography. 著者: Pavel V Afonine / Billy K Poon / Randy J Read / Oleg V Sobolev / Thomas C Terwilliger / Alexandre Urzhumtsev / Paul D Adams /   要旨: This article describes the implementation of real-space refinement in the phenix.real_space_refine program from the PHENIX suite. The use of a simplified refinement target function enables very fast ...This article describes the implementation of real-space refinement in the phenix.real_space_refine program from the PHENIX suite. The use of a simplified refinement target function enables very fast calculation, which in turn makes it possible to identify optimal data-restraint weights as part of routine refinements with little runtime cost. Refinement of atomic models against low-resolution data benefits from the inclusion of as much additional information as is available. In addition to standard restraints on covalent geometry, phenix.real_space_refine makes use of extra information such as secondary-structure and rotamer-specific restraints, as well as restraints or constraints on internal molecular symmetry. The re-refinement of 385 cryo-EM-derived models available in the Protein Data Bank at resolutions of 6 Å or better shows significant improvement of the models and of the fit of these models to the target maps. #2: ジャーナル: Nat Commun / 年: 2023タイトル: Structural basis of the mycobacterial stress-response RNA polymerase auto-inhibition via oligomerization. 著者: Zakia Morichaud / Stefano Trapani / Rishi K Vishwakarma / Laurent Chaloin / Corinne Lionne / Joséphine Lai-Kee-Him / Patrick Bron / Konstantin Brodolin / 要旨: Self-assembly of macromolecules into higher-order symmetric structures is fundamental for the regulation of biological processes. Higher-order symmetric structure self-assembly by the gene expression ...Self-assembly of macromolecules into higher-order symmetric structures is fundamental for the regulation of biological processes. Higher-order symmetric structure self-assembly by the gene expression machinery, such as bacterial DNA-dependent RNA polymerase (RNAP), has never been reported before. Here, we show that the stress-response σ factor from the human pathogen, Mycobacterium tuberculosis, induces the RNAP holoenzyme oligomerization into a supramolecular complex composed of eight RNAP units. Cryo-electron microscopy revealed a pseudo-symmetric structure of the RNAP octamer in which RNAP protomers are captured in an auto-inhibited state and display an open-clamp conformation. The structure shows that σ is sequestered by the RNAP flap and clamp domains. The transcriptional activator RbpA prevented octamer formation by promoting the initiation-competent RNAP conformation. Our results reveal that a non-conserved region of σ is an allosteric controller of transcription initiation and demonstrate how basal transcription factors can regulate gene expression by modulating the RNAP holoenzyme assembly and hibernation. #3: ジャーナル: Nucleic Acids Res / 年: 2018 タイトル: RbpA relaxes promoter selectivity of M. tuberculosis RNA polymerase. 著者: Ayyappasamy Sudalaiyadum Perumal / Rishi Kishore Vishwakarma / Yangbo Hu / Zakia Morichaud / Konstantin Brodolin /  要旨: The transcriptional activator RbpA associates with Mycobacterium tuberculosis RNA polymerase (MtbRNAP) during transcription initiation, and stimulates formation of the MtbRNAP-promoter open complex ...The transcriptional activator RbpA associates with Mycobacterium tuberculosis RNA polymerase (MtbRNAP) during transcription initiation, and stimulates formation of the MtbRNAP-promoter open complex (RPo). Here, we explored the influence of promoter motifs on RbpA-mediated activation of MtbRNAP containing the stress-response σB subunit. We show that both the 'extended -10' promoter motif (T-17G-16T-15G-14) and RbpA stabilized RPo and allowed promoter opening at suboptimal temperatures. Furthermore, in the presence of the T-17G-16T-15G-14 motif, RbpA was dispensable for RNA synthesis initiation, while exerting a stabilization effect on RPo. On the other hand, RbpA compensated for the lack of sequence-specific interactions of domains 3 and 4 of σB with the extended -10 and the -35 motifs, respectively. Mutations of the positively charged residues K73, K74 and R79 in RbpA basic linker (BL) had little effect on RPo formation, but affected MtbRNAP capacity for de novo transcription initiation. We propose that RbpA stimulates transcription by strengthening the non-specific interaction of the σ subunit with promoter DNA upstream of the -10 element, and by indirectly optimizing MtbRNAP interaction with initiation substrates. Consequently, RbpA renders MtbRNAP promiscuous in promoter selection, thus compensating for the weak conservation of the -35 motif in mycobacteria. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 9fjs.cif.gz | 592.4 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb9fjs.ent.gz | 463.3 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 9fjs.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/fj/9fjsftp://data.pdbj.org/pub/pdb/validation_reports/fj/9fjs | HTTPS FTP |
|---|
-関連構造データ







-リンク
 PDBj
PDBj






































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-DNA-directed RNA polymerase subunit ... , 4種, 5分子 abcde
| #1: タンパク質 | 分子量: 37745.328 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Mycobacterium tuberculosis H37Rv (結核菌)遺伝子: rpoA, Rv3457c, MTCY13E12.10c / プラスミド: pMR4 / 発現宿主: #2: タンパク質 | | 分子量: 129602.344 Da / 分子数: 1 / Mutation: L2E3G4C5I6 -> V / 由来タイプ: 組換発現 由来: (組換発現) Mycobacterium tuberculosis H37Rv (結核菌)遺伝子: rpoB, Rv0667, MTCI376.08c / プラスミド: pMR4 / 発現宿主: #3: タンパク質 | | 分子量: 147438.344 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) Mycobacterium tuberculosis H37Rv (結核菌)遺伝子: rpoC, Rv0668, MTCI376.07c / プラスミド: pMR4 / 発現宿主: #4: タンパク質 | | 分子量: 11851.140 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) Mycobacterium tuberculosis H37Rv (結核菌)遺伝子: rpoZ, Rv1390, MTCY21B4.07 / プラスミド: pMR4 / 発現宿主: |
|---|
-タンパク質 / DNA鎖 , 2種, 2分子 fO
| #5: タンパク質 | 分子量: 38572.773 Da / 分子数: 1 / 由来タイプ: 組換発現 詳細: Addition of twenty N-terminal residues (MGSSHHHHHHSSGLVPRGSH) including 6xHIS tag 由来: (組換発現) Mycobacterium tuberculosis H37Rv (結核菌)遺伝子: sigB, mysB, Rv2710 / プラスミド: pET28a / 詳細 (発現宿主): derivative comprising Rv2710 gene / 発現宿主: |
|---|---|
| #6: DNA鎖 | 分子量: 5297.444 Da / 分子数: 1 / 由来タイプ: 合成 / 詳細: synthetic oligonucleotide / 由来: (合成) |
-非ポリマー , 2種, 3分子 

| #7: 化合物 |  分子量: 65.409 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Zn#8: 化合物 | ChemComp-MG / |  分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION 分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|---|
| Has protein modification | N |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: RNA polymerase holoenzyme bound to -10 promoter element ssDNA oligo タイプ: COMPLEX / Entity ID: #1-#6 / 由来: RECOMBINANT | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: 0.406 MDa / 実験値: NO | |||||||||||||||||||||||||
| 由来(天然) | 生物種: Mycobacterium tuberculosis H37Rv (結核菌) | |||||||||||||||||||||||||
| 由来(組換発現) | 生物種: | |||||||||||||||||||||||||
| 緩衝液 | pH: 7.9 / 詳細: 8 mM CHAPSO added before vitrificaion | |||||||||||||||||||||||||
| 緩衝液成分 |
| |||||||||||||||||||||||||
| 試料 | 濃度: 6 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | |||||||||||||||||||||||||
| 試料支持 | グリッドの材料: GOLD / グリッドのサイズ: 200 divisions/in. / グリッドのタイプ: Quantifoil R2/2 | |||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 90 % / 凍結前の試料温度: 291.15 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 81000 X / 最大 デフォーカス(公称値): 2500 nm / 最小 デフォーカス(公称値): 800 nm / Cs: 0.01 mm / C2レンズ絞り径: 50 µm |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 平均露光時間: 1.997 sec. / 電子線照射量: 55.735 e/Å2 / フィルム・検出器のモデル: GATAN K3 (6k x 4k) / 撮影したグリッド数: 1 / 実像数: 9202 / 詳細: images were collected in super-resolution mode |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 695496 | ||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.48 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 67957 / アルゴリズム: FOURIER SPACE / クラス平均像の数: 1 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT / 空間: REAL | ||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | 3D fitting-ID: 1
| ||||||||||||||||||||||||||||||||||||||||
| 精密化 | 交差検証法: NONE |

