 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 9ff7 | ||||||
|---|---|---|---|---|---|---|---|
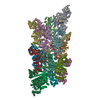
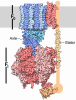
| タイトル | Structure of the BMOE-crosslinked transcription termination factor Rho in the presence of ppGpp; S84C/M405C double mutant | ||||||
 要素 要素 | Transcription termination factor Rho | ||||||
 キーワード キーワード | TRANSCRIPTION / Rho / termination / bacterial stress response / ppGpp | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報ATP-dependent activity, acting on RNA / helicase activity / DNA-templated transcription termination / 加水分解酵素; 酸無水物に作用; 酸無水物に作用・細胞または細胞小器官の運動に関与 / hydrolase activity / RNA binding / ATP binding / cytosol 類似検索 - 分子機能 | ||||||
| 生物種 |  | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.4 Å | ||||||
 データ登録者 データ登録者 | Said, N. / Hilal, T. / Wahl, M.C. | ||||||
| 資金援助 |  ドイツ, 1件 ドイツ, 1件
| ||||||
 引用 引用 | ジャーナル: Nat Commun / 年: 2025 タイトル: Nucleotide-induced hyper-oligomerization inactivates transcription termination factor ρ. 著者: Bing Wang / Nelly Said / Tarek Hilal / Mark Finazzo / Markus C Wahl / Irina Artsimovitch /  要旨: Bacterial RNA helicase ρ is a genome sentinel that terminates the synthesis of damaged and junk RNAs that are not translated by the ribosome. It is unclear how ρ is regulated during dormancy or ...Bacterial RNA helicase ρ is a genome sentinel that terminates the synthesis of damaged and junk RNAs that are not translated by the ribosome. It is unclear how ρ is regulated during dormancy or stress, when translation is inefficient and RNAs are vulnerable to ρ-mediated release. We use cryogenic electron microscopy, biochemical, and genetic approaches to show that substitutions of residues in the connector between two ρ domains or ADP promote the formation of extended Escherichia coli ρ filaments. By contrast, (p)ppGpp induces the formation of transient ρ dodecamers. Our results demonstrate that ADP and (p)ppGpp nucleotides bound at subunit interfaces inhibit ρ ring closure that underpins the hexamer activation, thus favoring the assembly of inactive higher-order oligomers. Connector substitutions and antibiotics that inhibit RNA and protein syntheses trigger ρ aggregation in the cell. These and other recent data implicate aggregation as a widespread strategy to tune ρ activity. #1: ジャーナル: bioRxiv / 年: 2023タイトル: Transcription termination factor ρ polymerizes under stress. 著者: Bing Wang / Nelly Said / Tarek Hilal / Mark Finazzo / Markus C Wahl / Irina Artsimovitch / 要旨: Bacterial RNA helicase ρ is a genome sentinel that terminates synthesis of damaged and junk RNAs that are not translated by the ribosome. Co-transcriptional RNA surveillance by ρ is essential for ...Bacterial RNA helicase ρ is a genome sentinel that terminates synthesis of damaged and junk RNAs that are not translated by the ribosome. Co-transcriptional RNA surveillance by ρ is essential for quality control of the transcriptome during optimal growth. However, it is unclear how bacteria protect their RNAs from overzealous ρ during dormancy or stress, conditions common in natural habitats. Here we used cryogenic electron microscopy, biochemical, and genetic approaches to show that residue substitutions, ADP, or ppGpp promote hyper-oligomerization of ρ. Our results demonstrate that nucleotides bound at subunit interfaces control ρ switching from active hexamers to inactive higher-order oligomers and extended filaments. Polymers formed upon exposure to antibiotics or ppGpp disassemble when stress is relieved, thereby directly linking termination activity to cellular physiology. Inactivation of ρ through hyper-oligomerization is a regulatory strategy shared by RNA polymerases, ribosomes, and metabolic enzymes across all life. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 9ff7.cif.gz | 831.5 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb9ff7.ent.gz | 705.3 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 9ff7.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/ff/9ff7ftp://data.pdbj.org/pub/pdb/validation_reports/ff/9ff7 | HTTPS FTP |
|---|
-関連構造データ

-リンク
 PDBj
PDBj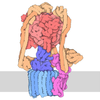

- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
| #1: タンパク質 | 分子量: 46971.098 Da / 分子数: 12 / 由来タイプ: 組換発現 / 由来: (組換発現) 参照: UniProt: P0AG32, 加水分解酵素; 酸無水物に作用; 酸無水物に作用・細胞または細胞小器官の運動に関与 #2: 化合物 | ChemComp-ME7 / 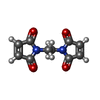  分子量: 220.182 Da / 分子数: 5 / 由来タイプ: 合成 / 式: C10H8N2O4 分子量: 220.182 Da / 分子数: 5 / 由来タイプ: 合成 / 式: C10H8N2O4#3: 化合物 | ChemComp-MG /   分子量: 24.305 Da / 分子数: 4 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 4 / 由来タイプ: 合成 / 式: Mg研究の焦点であるリガンドがあるか | N | Has protein modification | N | |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Dimer of transcription termination factor Rho / タイプ: COMPLEX / 詳細: S84C/M405C double mutant, BMOE crosslinked / Entity ID: #1 / 由来: RECOMBINANT |
|---|---|
| 由来(天然) | 生物種: |
| 由来(組換発現) | 生物種: |
| 緩衝液 | pH: 8 |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 試料支持 | グリッドの材料: COPPER / グリッドのサイズ: 200 divisions/in. / グリッドのタイプ: Quantifoil R1.2/1.3 |
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 283 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 96000 X / 最大 デフォーカス(公称値): 2000 nm / 最小 デフォーカス(公称値): 900 nm / Cs: 2.7 mm / C2レンズ絞り径: 50 µm / アライメント法: ZEMLIN TABLEAU |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 平均露光時間: 40.57 sec. / 電子線照射量: 44 e/Å2 / 検出モード: COUNTING フィルム・検出器のモデル: FEI FALCON III (4k x 4k) 撮影したグリッド数: 1 / 実像数: 2218 |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 840400 | ||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.4 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 96116 / アルゴリズム: BACK PROJECTION / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT / 空間: REAL | ||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|
