 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-9dul: Structure of mutant 30S subunit with extended helix 26, version 4 -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 9dul | ||||||
|---|---|---|---|---|---|---|---|
| Title | Structure of mutant 30S subunit with extended helix 26, version 4 | ||||||
 Components Components |
| ||||||
 Keywords Keywords | RIBOSOME / Translation / 30S subunit / rRNA / Ribosomal Proteins | ||||||
| Function / homology |  Function and homology information Function and homology informationnegative regulation of translational initiation / mRNA regulatory element binding translation repressor activity / transcription antitermination / DNA-templated transcription termination / maintenance of translational fidelity / mRNA 5'-UTR binding / ribosomal small subunit assembly / ribosomal small subunit biogenesis / small ribosomal subunit / small ribosomal subunit rRNA binding ...negative regulation of translational initiation / mRNA regulatory element binding translation repressor activity / transcription antitermination / DNA-templated transcription termination / maintenance of translational fidelity / mRNA 5'-UTR binding / ribosomal small subunit assembly / ribosomal small subunit biogenesis / small ribosomal subunit / small ribosomal subunit rRNA binding / cytosolic small ribosomal subunit / cytoplasmic translation / tRNA binding / negative regulation of translation / rRNA binding / structural constituent of ribosome / ribosome / translation / ribonucleoprotein complex / response to antibiotic / mRNA binding / RNA binding / membrane / cytoplasm / cytosol Similarity search - Function | ||||||
| Biological species |  | ||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 2.56 Å | ||||||
 Authors Authors | Boyko, K. / Cate, J. | ||||||
| Funding support |  United States, 1items United States, 1items
| ||||||
 Citation Citation | Journal: Biochemistry / Year: 2025 Title: Role of Ribosomal Protein bS1 in Orthogonal mRNA Start Codon Selection. Authors: Kristina V Boyko / Rebecca A Bernstein / Minji Kim / Jamie H D Cate / Abstract: In many bacteria, the location of the mRNA start codon is determined by a short ribosome binding site sequence that base pairs with the 3'-end of 16S rRNA (rRNA) in the 30S subunit. Many groups have ...In many bacteria, the location of the mRNA start codon is determined by a short ribosome binding site sequence that base pairs with the 3'-end of 16S rRNA (rRNA) in the 30S subunit. Many groups have changed these short sequences, termed the Shine-Dalgarno (SD) sequence in the mRNA and the anti-Shine-Dalgarno (ASD) sequence in 16S rRNA, to create "orthogonal" ribosomes to enable the synthesis of orthogonal polymers in the presence of the endogenous translation machinery. However, orthogonal ribosomes are prone to SD-independent translation. Ribosomal protein bS1, which binds to the 30S ribosomal subunit, is thought to promote translation initiation by shuttling the mRNA to the ribosome. Thus, a better understanding of how the SD and bS1 contribute to start codon selection could help efforts to improve the orthogonality of ribosomes. Here, we engineered the ribosome to prevent binding of bS1 to the 30S subunit and separate the activity of bS1 binding to the ribosome from the role of the mRNA SD sequence in start codon selection. We find that ribosomes lacking bS1 are slightly less active than wild-type ribosomes in vitro. Furthermore, orthogonal 30S subunits lacking bS1 do not have an improved orthogonality. Our findings suggest that mRNA features outside the SD sequence and independent of binding of bS1 to the ribosome likely contribute to start codon selection and the lack of orthogonality of present orthogonal ribosomes. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 9dul.cif.gz | 1.2 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb9dul.ent.gz | 956.4 KB | Display | PDB format |
| PDBx/mmJSON format | 9dul.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/du/9dulftp://data.pdbj.org/pub/pdb/validation_reports/du/9dul | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  47169MC  9dukC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |

-Links
 PDBj
PDBj





























- Assembly
Assembly
| Deposited unit | 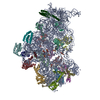
|
|---|---|
| 1 |
|
-Components
-RNA chain , 1 types, 1 molecules A
| #1: RNA chain | Mass: 496993.688 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|
-Small ribosomal subunit protein ... , 20 types, 20 molecules BCDEFGHIJKLMNOPQRSTU
| #2: Protein | Mass: 25989.867 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|---|
| #3: Protein | Mass: 26031.316 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #4: Protein | Mass: 23514.199 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #5: Protein | Mass: 17629.398 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #6: Protein | Mass: 15727.512 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #7: Protein | Mass: 20055.156 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #8: Protein | Mass: 14146.557 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #9: Protein | Mass: 14886.270 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #10: Protein | Mass: 11755.597 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #11: Protein | Mass: 13871.959 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #12: Protein | Mass: 13814.249 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #13: Protein | Mass: 13128.467 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #14: Protein | Mass: 11606.560 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #15: Protein | Mass: 10290.816 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #16: Protein | Mass: 9207.572 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #17: Protein | Mass: 9724.491 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #18: Protein | Mass: 9005.472 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #19: Protein | Mass: 10455.355 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #20: Protein | Mass: 9708.464 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #21: Protein | Mass: 8524.039 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
-Details
| Has ligand of interest | Y |
|---|---|
| Has protein modification | Y |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: 30S subunit / Type: RIBOSOME / Entity ID: all / Source: NATURAL |
|---|---|
| Molecular weight | Experimental value: NO |
| Source (natural) | Organism: |
| Buffer solution | pH: 7.5 |
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Specimen support | Grid type: Quantifoil R1.2/1.3 |
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: TFS KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal defocus max: 1500 nm / Nominal defocus min: 500 nm |
| Image recording | Electron dose: 40 e/Å2 / Film or detector model: GATAN K3 (6k x 4k) |
- Processing
Processing
| EM software | Name: PHENIX / Version: 1.20_4459: / Category: model refinement |
|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION |
| 3D reconstruction | Resolution: 2.56 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 51527 / Symmetry type: POINT |
