ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
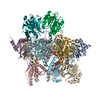
| 登録情報 | データベース: PDB / ID: 9d0t | ||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Proteasome core particle assembly intermediate Blm10:13S purified from Saccharomyces cerevisiae | ||||||||||||||||||||||||||||||||||||||||||||||||
 要素 要素 |
| ||||||||||||||||||||||||||||||||||||||||||||||||
 キーワード キーワード | HYDROLASE / Proteasome / assembly chaperone / Blm10 / PA200 / Blm10-13S / Pba1 / CP | ||||||||||||||||||||||||||||||||||||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報proteasome core complex import into nucleus / myosin-light-chain kinase / myosin light chain kinase activity / myosin light chain binding / proteasome storage granule assembly / cardiac muscle tissue morphogenesis / peptidase activator activity / proteasome core complex assembly / nuclear outer membrane-endoplasmic reticulum membrane network / Proteasome assembly ...proteasome core complex import into nucleus / myosin-light-chain kinase / myosin light chain kinase activity / myosin light chain binding / proteasome storage granule assembly / cardiac muscle tissue morphogenesis / peptidase activator activity / proteasome core complex assembly / nuclear outer membrane-endoplasmic reticulum membrane network / Proteasome assembly / Cross-presentation of soluble exogenous antigens (endosomes) / TNFR2 non-canonical NF-kB pathway / Ubiquitin-Mediated Degradation of Phosphorylated Cdc25A / proteasomal ubiquitin-independent protein catabolic process / Regulation of PTEN stability and activity / CDK-mediated phosphorylation and removal of Cdc6 / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / KEAP1-NFE2L2 pathway / Neddylation / proteasome binding / Orc1 removal from chromatin / MAPK6/MAPK4 signaling / proteasome storage granule / Antigen processing: Ubiquitination & Proteasome degradation / Ub-specific processing proteases / proteasome endopeptidase complex / endopeptidase activator activity / proteasome core complex, beta-subunit complex / proteasome assembly / threonine-type endopeptidase activity / proteasome core complex, alpha-subunit complex / striated muscle contraction / : / regulation of proteasomal protein catabolic process / Neutrophil degranulation / proteasome complex / ubiquitin-dependent protein catabolic process / endopeptidase activity / proteasome-mediated ubiquitin-dependent protein catabolic process / calmodulin binding / DNA repair / mRNA binding / DNA damage response / endoplasmic reticulum membrane / signal transduction / mitochondrion / ATP binding / nucleus / cytoplasm / cytosol 類似検索 - 分子機能 | ||||||||||||||||||||||||||||||||||||||||||||||||
| 生物種 |  | ||||||||||||||||||||||||||||||||||||||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 2.84 Å | ||||||||||||||||||||||||||||||||||||||||||||||||
 データ登録者 データ登録者 | Chen, X. / Kaur, M. / Roelofs, J. / Walters, K.J. | ||||||||||||||||||||||||||||||||||||||||||||||||
| 資金援助 |  米国, 2件 米国, 2件
| ||||||||||||||||||||||||||||||||||||||||||||||||
 引用 引用 | ジャーナル: bioRxiv / 年: 2024 タイトル: Structure of Blm10:13S proteasome intermediate reveals parallel assembly pathways for the proteasome core particle. 著者: Mandeep Kaur / Xiang Chen / Stella Y Lee / Tyler M Weaver / Bret D Freudenthal / Kylie J Walters / Jeroen Roelofs / 要旨: Proteasomes are formed by chaperone-assisted assembly of core particles (CPs) and regulatory particles (RPs). The CP chaperone dimer Pba1/Pba2 binds early to proteasome subunits, and is thought to be ...Proteasomes are formed by chaperone-assisted assembly of core particles (CPs) and regulatory particles (RPs). The CP chaperone dimer Pba1/Pba2 binds early to proteasome subunits, and is thought to be replaced by Blm10 to form Blm10:CP, which promotes ATP-independent degradation of disordered proteins. Here, we present evidence of distinct parallel assembly pathways for CP by solving five cryo-EM structures including a Blm10:13S pre-assembly intermediate. Our data conflict with the current model of Blm10 and Pba1/Pba2 sequential activity in a single assembly pathway, as we find their CP binding is mutually exclusive and both are present on early and late assembly intermediates. CP affinity for Pba1/Pba2 is reduced during maturation, promoting Pba1/Pba2 release. We find Blm10 undergoes no such affinity switch, suggesting this pathway predominantly yields mature Blm10-bound CP. Altogether, our findings conflict with the current paradigm of sequential CP binding to instead indicate parallel assembly pathways by Pba1/Pba2 and Blm10. | ||||||||||||||||||||||||||||||||||||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 9d0t.cif.gz | 1.5 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb9d0t.ent.gz | 1.2 MB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 9d0t.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 9d0t_validation.pdf.gz | 1.2 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 9d0t_full_validation.pdf.gz | 1.2 MB | 表示 | |
| XML形式データ | 9d0t_validation.xml.gz | 107.3 KB | 表示 | |
| CIF形式データ | 9d0t_validation.cif.gz | 176.8 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/d0/9d0tftp://data.pdbj.org/pub/pdb/validation_reports/d0/9d0t | HTTPS FTP |
-関連構造データ




-リンク
 PDBj
PDBj













- 集合体
集合体
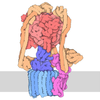
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-Proteasome subunit alpha type- ... , 6種, 6分子 ABCDEF
| #1: タンパク質 | 分子量: 28033.830 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: SCL1, PRC2, PRS2, YGL011C / 発現宿主: |
|---|---|
| #2: タンパク質 | 分子量: 27191.828 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: PRE8, PRS4, YML092C / 発現宿主: |
| #3: タンパク質 | 分子量: 28748.230 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: PRE9, PRS5, YGR135W / 発現宿主: |
| #4: タンパク質 | 分子量: 28478.111 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: PRE6, YOL038W / 発現宿主: |
| #5: タンパク質 | 分子量: 28649.086 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: PUP2, DOA5, YGR253C, G9155 / 発現宿主: |
| #6: タンパク質 | 分子量: 25634.000 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: PRE5, YMR314W, YM9924.06 / 発現宿主: |
-タンパク質 , 3種, 3分子 GOP
| #7: タンパク質 | 分子量: 31575.068 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: PRE10, PRC1, PRS1, YOR362C, O6650 / 発現宿主: |
|---|---|
| #11: タンパク質 | 分子量: 246282.375 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: BLM10, BLM3, YFL007W, YFL006W / 発現宿主: |
| #12: タンパク質 | 分子量: 22753.646 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: UMP1, YBR173C, YBR1234, MYLK2 / 発現宿主: 参照: UniProt: P38293, UniProt: A4IFM7, myosin-light-chain kinase |
-Proteasome subunit beta type- ... , 3種, 3分子 IJK
| #8: タンパク質 | 分子量: 28299.889 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: PUP1, YOR157C / 発現宿主: |
|---|---|
| #9: タンパク質 | 分子量: 22627.842 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: PUP3, YER094C / 発現宿主: |
| #10: タンパク質 | 分子量: 22545.676 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: PRE1, YER012W / 発現宿主: |
-詳細
| Has protein modification | N |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Proteasome assembly intermediate / タイプ: COMPLEX / Entity ID: all / 由来: RECOMBINANT | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: 0.534 MDa / 実験値: NO | |||||||||||||||||||||||||
| 由来(天然) | 生物種: | |||||||||||||||||||||||||
| 由来(組換発現) | 生物種: | |||||||||||||||||||||||||
| 緩衝液 | pH: 7.5 | |||||||||||||||||||||||||
| 緩衝液成分 |
| |||||||||||||||||||||||||
| 試料 | 濃度: 1.8 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | |||||||||||||||||||||||||
| 試料支持 | 詳細: 0.42 mBar, 15mA / グリッドの材料: COPPER / グリッドのサイズ: 400 divisions/in. / グリッドのタイプ: Quantifoil R1.2/1.3 | |||||||||||||||||||||||||
| 急速凍結 | 装置: LEICA EM GP / 凍結剤: ETHANE / 湿度: 80 % / 凍結前の試料温度: 298.15 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: TFS KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 81000 X / 最大 デフォーカス(公称値): 2500 nm / 最小 デフォーカス(公称値): 900 nm / Cs: 2.7 mm |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 電子線照射量: 60 e/Å2 フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) 実像数: 5235 |
| 電子光学装置 | エネルギーフィルター名称: GIF Bioquantum |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 4058518 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 2.84 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 573652 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | B value: 120.13 / プロトコル: RIGID BODY FIT / 空間: REAL / Target criteria: Cross-correlation coefficient | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | 3D fitting-ID: 1 / Source name: PDB / タイプ: experimental model
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 | 最高解像度: 2.84 Å |