 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 8xgc | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Structure of yeast replisome associated with FACT and histone hexamer, Composite map | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | REPLICATION / Replisome / FACT / histone hexamer | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報Regulation of TP53 Activity through Phosphorylation / : / regulation of sister chromatid cohesion / establishment of sister chromatid cohesion / DNA-templated DNA replication maintenance of fidelity / gene conversion / FACT complex / Unwinding of DNA / maintenance of DNA repeat elements / regulation of nuclear cell cycle DNA replication ...Regulation of TP53 Activity through Phosphorylation / : / regulation of sister chromatid cohesion / establishment of sister chromatid cohesion / DNA-templated DNA replication maintenance of fidelity / gene conversion / FACT complex / Unwinding of DNA / maintenance of DNA repeat elements / regulation of nuclear cell cycle DNA replication / DNA replication initiation / replication fork arrest / Cul8-RING ubiquitin ligase complex / meiotic chromosome segregation / HATs acetylate histones / RNA polymerase I upstream activating factor complex / Condensation of Prophase Chromosomes / SIRT1 negatively regulates rRNA expression / epsilon DNA polymerase complex / MCM core complex / Assembly of the pre-replicative complex / Switching of origins to a post-replicative state / DNA strand elongation involved in mitotic DNA replication / GINS complex / MCM complex binding / mitotic DNA replication preinitiation complex assembly / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / nuclear DNA replication / regulation of chromatin organization / premeiotic DNA replication / Assembly of the ORC complex at the origin of replication / replication fork protection complex / HDACs deacetylate histones / pre-replicative complex assembly involved in nuclear cell cycle DNA replication / SUMO binding / anaphase-promoting complex binding / nucleotide-excision repair, DNA gap filling / Activation of the pre-replicative complex / mitotic DNA replication / DNA replication checkpoint signaling / CMG complex / establishment of mitotic sister chromatid cohesion / nuclear pre-replicative complex / DNA replication proofreading / Termination of translesion DNA synthesis / Activation of ATR in response to replication stress / nucleosome organization / DNA replication preinitiation complex / single-stranded DNA 3'-5' DNA exonuclease activity / MCM complex / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / mitotic DNA replication checkpoint signaling / Oxidative Stress Induced Senescence / double-strand break repair via break-induced replication / mitotic DNA replication initiation / RMTs methylate histone arginines / cellular response to osmotic stress / single-stranded DNA helicase activity / mitotic intra-S DNA damage checkpoint signaling / SUMOylation of chromatin organization proteins / silent mating-type cassette heterochromatin formation / 加水分解酵素; エステル加水分解酵素; 5'-リン酸モノエステル産生エンドデオキシリボヌクレアーゼ / regulation of DNA-templated DNA replication initiation / mitotic sister chromatid cohesion / TP53 Regulates Transcription of DNA Repair Genes / RNA Polymerase II Pre-transcription Events / DNA strand elongation involved in DNA replication / nuclear chromosome / 3'-5' DNA helicase activity / leading strand elongation / RNA Polymerase I Promoter Escape / positive regulation of transcription by RNA polymerase I / mitotic G2 DNA damage checkpoint signaling / nucleolar large rRNA transcription by RNA polymerase I / nuclear replication fork / replication fork processing / Estrogen-dependent gene expression / rRNA transcription / DNA replication origin binding / Dual incision in TC-NER / positive regulation of RNA polymerase II transcription preinitiation complex assembly / DNA replication initiation / positive regulation of transcription initiation by RNA polymerase II / Ub-specific processing proteases / error-prone translesion synthesis / subtelomeric heterochromatin formation / base-excision repair, gap-filling / telomere maintenance / nuclear periphery / DNA helicase activity / replication fork / meiotic cell cycle / transcription elongation by RNA polymerase II / helicase activity / base-excision repair / double-strand break repair via nonhomologous end joining / DNA-templated DNA replication / structural constituent of chromatin / nucleosome / heterochromatin formation 類似検索 - 分子機能 | ||||||
| 生物種 |  | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.7 Å | ||||||
 データ登録者 データ登録者 | Li, N. / Gao, Y. / Yu, D. / Gao, N. / Zhai, Y. | ||||||
| 資金援助 |  中国, 1件 中国, 1件
| ||||||
 引用 引用 | ジャーナル: Nature / 年: 2024 タイトル: Parental histone transfer caught at the replication fork. 著者: Ningning Li / Yuan Gao / Yujie Zhang / Daqi Yu / Jianwei Lin / Jianxun Feng / Jian Li / Zhichun Xu / Yingyi Zhang / Shangyu Dang / Keda Zhou / Yang Liu / Xiang David Li / Bik Kwoon Tye / Qing ...著者: Ningning Li / Yuan Gao / Yujie Zhang / Daqi Yu / Jianwei Lin / Jianxun Feng / Jian Li / Zhichun Xu / Yingyi Zhang / Shangyu Dang / Keda Zhou / Yang Liu / Xiang David Li / Bik Kwoon Tye / Qing Li / Ning Gao / Yuanliang Zhai /  要旨: In eukaryotes, DNA compacts into chromatin through nucleosomes. Replication of the eukaryotic genome must be coupled to the transmission of the epigenome encoded in the chromatin. Here we report cryo- ...In eukaryotes, DNA compacts into chromatin through nucleosomes. Replication of the eukaryotic genome must be coupled to the transmission of the epigenome encoded in the chromatin. Here we report cryo-electron microscopy structures of yeast (Saccharomyces cerevisiae) replisomes associated with the FACT (facilitates chromatin transactions) complex (comprising Spt16 and Pob3) and an evicted histone hexamer. In these structures, FACT is positioned at the front end of the replisome by engaging with the parental DNA duplex to capture the histones through the middle domain and the acidic carboxyl-terminal domain of Spt16. The H2A-H2B dimer chaperoned by the carboxyl-terminal domain of Spt16 is stably tethered to the H3-H4 tetramer, while the vacant H2A-H2B site is occupied by the histone-binding domain of Mcm2. The Mcm2 histone-binding domain wraps around the DNA-binding surface of one H3-H4 dimer and extends across the tetramerization interface of the H3-H4 tetramer to the binding site of Spt16 middle domain before becoming disordered. This arrangement leaves the remaining DNA-binding surface of the other H3-H4 dimer exposed to additional interactions for further processing. The Mcm2 histone-binding domain and its downstream linker region are nested on top of Tof1, relocating the parental histones to the replisome front for transfer to the newly synthesized lagging-strand DNA. Our findings offer crucial structural insights into the mechanism of replication-coupled histone recycling for maintaining epigenetic inheritance. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8xgc.cif.gz | 1.9 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8xgc.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 8xgc.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 8xgc_validation.pdf.gz | 1.4 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 8xgc_full_validation.pdf.gz | 1.4 MB | 表示 | |
| XML形式データ | 8xgc_validation.xml.gz | 243.6 KB | 表示 | |
| CIF形式データ | 8xgc_validation.cif.gz | 379.6 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/xg/8xgcftp://data.pdbj.org/pub/pdb/validation_reports/xg/8xgc | HTTPS FTP |
-関連構造データ





-リンク
 PDBj
PDBj







































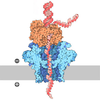





- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-DNA replication licensing factor ... , 5種, 5分子 23467
| #1: タンパク質 | 分子量: 98911.539 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #2: タンパク質 | 分子量: 107653.508 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #3: タンパク質 | 分子量: 105138.375 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #5: タンパク質 | 分子量: 113110.211 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #6: タンパク質 | 分子量: 95049.875 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-タンパク質 , 10種, 14分子 5EFGHIJKNROSPQ
| #4: タンパク質 | 分子量: 86505.734 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| #13: タンパク質 | 分子量: 74324.836 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) | ||||||||||||||
| #14: タンパク質 | 分子量: 104543.391 Da / 分子数: 3 / 由来タイプ: 天然 / 由来: (天然) #15: タンパク質 | | 分子量: 141296.875 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) #16: タンパク質 | | 分子量: 36402.590 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) #17: タンパク質 | | 分子量: 124516.375 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) #20: タンパク質 | 分子量: 15391.007 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #21: タンパク質 | 分子量: 11395.390 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #22: タンパク質 | | 分子量: 14013.177 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) #23: タンパク質 | | 分子量: 14264.341 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-DNA polymerase epsilon ... , 2種, 2分子 89
| #7: タンパク質 | 分子量: 255992.484 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) 参照: UniProt: P21951, DNA-directed DNA polymerase, 加水分解酵素; エステル加水分解酵素; 5'-リン酸モノエステル産生エンドデオキシリボヌクレアーゼ |
|---|---|
| #8: タンパク質 | 分子量: 78425.852 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-DNA replication complex GINS protein ... , 4種, 4分子 ABCD
| #9: タンパク質 | 分子量: 24230.576 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #10: タンパク質 | 分子量: 29341.074 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #11: タンパク質 | 分子量: 21977.135 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #12: タンパク質 | 分子量: 33983.617 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-FACT complex subunit ... , 2種, 2分子 LM
| #18: タンパク質 | 分子量: 118776.984 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #19: タンパク質 | 分子量: 63068.594 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-DNA鎖 , 2種, 2分子 XY
| #24: DNA鎖 | 分子量: 15689.072 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #25: DNA鎖 | 分子量: 11935.764 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-非ポリマー , 2種, 12分子 

| #26: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 7 / 由来タイプ: 合成 / 式: Zn / タイプ: SUBJECT OF INVESTIGATION 分子量: 65.409 Da / 分子数: 7 / 由来タイプ: 合成 / 式: Zn / タイプ: SUBJECT OF INVESTIGATION#27: 化合物 | ChemComp-ADP /  分子量: 427.201 Da / 分子数: 5 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / タイプ: SUBJECT OF INVESTIGATION / コメント: ADP, エネルギー貯蔵分子*YM 分子量: 427.201 Da / 分子数: 5 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / タイプ: SUBJECT OF INVESTIGATION / コメント: ADP, エネルギー貯蔵分子*YM |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|---|
| Has protein modification | Y |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Endogenous replisomes / タイプ: COMPLEX / Entity ID: #1-#25 / 由来: NATURAL |
|---|---|
| 由来(天然) | 生物種: |
| 緩衝液 | pH: 7.5 |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 急速凍結 | 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 最大 デフォーカス(公称値): 2500 nm / 最小 デフォーカス(公称値): 1000 nm |
| 撮影 | 電子線照射量: 50 e/Å2 フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) |
- 解析
解析
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION |
|---|---|
| 3次元再構成 | 解像度: 3.7 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 524000 / 対称性のタイプ: POINT |
