 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8vjm | |||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | SpoIVFB(E44Q variant):pro-sigmaK complex | |||||||||||||||||||||||||||||||||||||||||||||
 Components Components |
| |||||||||||||||||||||||||||||||||||||||||||||
 Keywords Keywords | MEMBRANE PROTEIN / spoIVFB / sigmaK / S2P / site-2-protease / protease | |||||||||||||||||||||||||||||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationHydrolases; Acting on peptide bonds (peptidases); Metalloendopeptidases / sigma factor antagonist complex / sporulation resulting in formation of a cellular spore / sigma factor activity / DNA-templated transcription initiation / metallopeptidase activity / DNA-directed RNA polymerase activity / transcription cis-regulatory region binding / regulation of DNA-templated transcription / proteolysis / metal ion binding Similarity search - Function | |||||||||||||||||||||||||||||||||||||||||||||
| Biological species |  | |||||||||||||||||||||||||||||||||||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 4 Å | |||||||||||||||||||||||||||||||||||||||||||||
 Authors Authors | Orlando, M.A. / Pouillon, H.J.T. / Mandal, S. / Kroos, L. / Orlando, B.J. | |||||||||||||||||||||||||||||||||||||||||||||
| Funding support |  United States, 2items United States, 2items
| |||||||||||||||||||||||||||||||||||||||||||||
 Citation Citation | Journal: Nat Commun / Year: 2024 Title: Substrate engagement by the intramembrane metalloprotease SpoIVFB. Authors: Melanie A Orlando / Hunter J T Pouillon / Saikat Mandal / Lee Kroos / Benjamin J Orlando / Abstract: S2P intramembrane metalloproteases regulate diverse signaling pathways across all three domains of life. However, the mechanism by which S2P metalloproteases engage substrates and catalyze peptide ...S2P intramembrane metalloproteases regulate diverse signaling pathways across all three domains of life. However, the mechanism by which S2P metalloproteases engage substrates and catalyze peptide hydrolysis within lipid membranes has remained elusive. Here we determine the cryo-EM structure of the S2P family intramembrane metalloprotease SpoIVFB from Bacillus subtilis bound to its native substrate Pro-σ. The structure and accompanying biochemical data demonstrate that SpoIVFB positions Pro-σ at the enzyme active site through a β-sheet augmentation mechanism, and reveal key interactions between Pro-σ and the interdomain linker connecting SpoIVFB transmembrane and CBS domains. The cryo-EM structure and molecular dynamics simulation reveal a plausible path for water to access the membrane-buried active site of SpoIVFB, and suggest a possible role of membrane lipids in facilitating substrate capture. These results provide key insight into how S2P intramembrane metalloproteases capture and position substrates for hydrolytic proteolysis within the hydrophobic interior of a lipid membrane. | |||||||||||||||||||||||||||||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8vjm.cif.gz | 161.1 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8vjm.ent.gz | 124.4 KB | Display | PDB format |
| PDBx/mmJSON format | 8vjm.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/vj/8vjmftp://data.pdbj.org/pub/pdb/validation_reports/vj/8vjm | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  43289MC  8vjlC C: citing same article ( M: map data used to model this data |
|---|---|
| Similar structure data |

-Links
 PDBj
PDBj

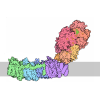

- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
| #1: Protein | Mass: 36990.527 Da / Num. of mol.: 2 / Mutation: E44Q Source method: isolated from a genetically manipulated source Source: (gene. exp.) Gene: spoIVFB, bofB, BSU27970 / Production host: References: UniProt: P26937, Hydrolases; Acting on peptide bonds (peptidases); Metalloendopeptidases #2: Protein | Mass: 15089.531 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Gene: sigK, cisB, spoIIIC, spoIVCB, BSU25760/BSU26390 / Production host: #3: Chemical |   Mass: 1005.188 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C47H88O22 / Comment: detergent*YM Mass: 1005.188 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C47H88O22 / Comment: detergent*YM#4: Chemical | ChemComp-PEF / |   Mass: 691.959 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C37H74NO8P / Comment: phospholipid*YM Mass: 691.959 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C37H74NO8P / Comment: phospholipid*YMHas ligand of interest | N | Has protein modification | N | |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: Complex of SpoIVFB (E44Q variant) and pro-sigmaK / Type: COMPLEX / Entity ID: #1-#2 / Source: RECOMBINANT | |||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Value: 0.052 MDa / Experimental value: NO | |||||||||||||||||||||||||||||||||||
| Source (natural) | Organism: | |||||||||||||||||||||||||||||||||||
| Source (recombinant) | Organism: | |||||||||||||||||||||||||||||||||||
| Buffer solution | pH: 8 Details: 25mM Tris, 150mM NaCl, 5mM betamercaptoethanol, 2mM MgCl2, 2mM ATP and 0.005% LMNG | |||||||||||||||||||||||||||||||||||
| Buffer component |
| |||||||||||||||||||||||||||||||||||
| Specimen | Conc.: 8 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES Details: Detergent solubilized complex of spoIVFB (E44Q variant) and pro-sigmaK(1-127) | |||||||||||||||||||||||||||||||||||
| Specimen support | Details: Glow-discharge in a Pelco EasyGlow at 15mA for 45s / Grid material: COPPER / Grid mesh size: 200 divisions/in. / Grid type: Quantifoil R2/2 | |||||||||||||||||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK IV / Cryogen name: ETHANE / Humidity: 100 % / Chamber temperature: 277 K |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: TFS KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 105000 X / Nominal defocus max: 2000 nm / Nominal defocus min: 800 nm / Cs: 2.7 mm / Alignment procedure: ZEMLIN TABLEAU |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Electron dose: 50 e/Å2 / Film or detector model: GATAN K3 (6k x 4k) |
| EM imaging optics | Energyfilter name: GIF Bioquantum / Energyfilter slit width: 20 eV |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| 3D reconstruction | Resolution: 4 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 78352 / Symmetry type: POINT | ||||||||||||||||||||||||
| Atomic model building | Space: REAL | ||||||||||||||||||||||||
| Refine LS restraints |
|
