プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray
放射波長
波長: 0.965459 Å / 相対比: 1
反射
解像度: 1.984→34.299 Å / Num. obs: 34279 / % possible obs: 96.6 % / 冗長度: 39.82 % 詳細: Some remarks regarding the mmCIF items written, the PDB Exchange Dictionary (PDBx/mmCIF) Version 5.0 supporting the data files in the current PDB archive (dictionary version 5.325, last ...詳細: Some remarks regarding the mmCIF items written, the PDB Exchange Dictionary (PDBx/mmCIF) Version 5.0 supporting the data files in the current PDB archive (dictionary version 5.325, last updated 2020-04-13: http://mmcif.wwpdb.org/dictionaries/mmcif_pdbx_v50.dic/Index/) and the actual quantities provided by MRFANA (https://github.com/githubgphl/MRFANA) from the autoPROC package (https://www.globalphasing.com/autoproc/). In general, the mmCIF categories here should provide items that are currently used in the PDB archive. If there are alternatives, the one recommended by the PDB developers has been selected. The distinction between *_all and *_obs quantities is not always clear: often only one version is actively used within the PDB archive (or is the one recommended by PDB developers). The intention of distinguishing between classes of reflections before and after some kind of observation criterion was applied, can in principle be useful - but such criteria change in various ways throughout the data processing steps (rejection of overloaded or too partial reflections, outlier/misfit rejections during scaling etc) and there is no retrospect computation of data scaling/merging statistics for the reflections used in the final refinement (where another observation criterion might have been applied). Typical data processing will usually only provide one version of statistics at various stages and these are given in the recommended item here, irrespective of the "_all" and "_obs" connotation, see e.g. the use of _reflns.pdbx_Rmerge_I_obs, _reflns.pdbx_Rrim_I_all and _reflns.pdbx_Rpim_I_all. Please note that all statistics related to "merged intensities" (or "merging") are based on inverse-variance weighting of the individual measurements making up a symmetry-unique reflection. This is standard for several decades now, even if some of the dictionary definitions seem to suggest that a simple "mean" or "average" intensity is being used instead. R-values are always given for all symmetry-equivalent reflections following Friedel's law, i.e. Bijvoet pairs are not treated separately (since we want to describe the overall mean intensity and not the mean I(+) and I(-) here). The Rrim metric is identical to the Rmeas R-value and only differs in name. _reflns.pdbx_number_measured_all is the number of measured intensities just before the final merging step (at which point no additional rejection takes place). _reflns.number_obs is the number of symmetry-unique observations, i.e. the result of merging those measurements via inverse-variance weighting. _reflns.pdbx_netI_over_sigmaI is based on the merged intensities (_reflns.number_obs) as expected. _reflns.pdbx_redundancy is synonymous with "multiplicity". The per-shell item _reflns_shell.number_measured_all corresponds to the overall value _reflns.pdbx_number_measured_all. The per-shell item _reflns_shell.number_unique_all corresponds to the overall value _reflns.number_obs. The per-shell item _reflns_shell.percent_possible_all corresponds to the overall value _reflns.percent_possible_obs. The per-shell item _reflns_shell.meanI_over_sigI_obs corresponds to the overall value given as _reflns.pdbx_netI_over_sigmaI. But be aware of the incorrect definition of the former in the current dictionary! CC1/2: 1 / CC1/2 anomalous: -0.372 / Rmerge(I) obs: 0.0874 / Rpim(I) all: 0.0141 / Rrim(I) all: 0.0886 / AbsDiff over sigma anomalous: 0.748 / Aniso diffraction limit 1: 1.984 Å / Aniso diffraction limit 2: 1.984 Å / Aniso diffraction limit 3: 1.984 Å / Aniso diffraction limit axis 1 ortho1: 1 / Aniso diffraction limit axis 1 ortho2: 0 / Aniso diffraction limit axis 1 ortho3: 0 / Aniso diffraction limit axis 2 ortho1: 0 / Aniso diffraction limit axis 2 ortho2: 1 / Aniso diffraction limit axis 2 ortho3: 0 / Aniso diffraction limit axis 3 ortho1: 0 / Aniso diffraction limit axis 3 ortho2: 0 / Aniso diffraction limit axis 3 ortho3: 1 / Net I/σ(I): 29.54 / Num. measured all: 1364986 / Observed signal threshold: 1.2 / Orthogonalization convention: pdb / % possible anomalous: 96.6 / % possible ellipsoidal: 96.6 / % possible ellipsoidal anomalous: 96.6 / % possible spherical: 96.6 / % possible spherical anomalous: 96.6 / Redundancy anomalous: 20.54 / Signal type: local
反射 シェル
解像度 (Å)
冗長度 (%)
Rmerge(I) obs
Mean I/σ(I) obs
Num. measured all
Num. measured obs
Num. unique all
Num. unique obs
CC1/2
CC1/2 anomalous
Rpim(I) all
Rrim(I) all
AbsDiff over sigma anomalous
% possible anomalous
% possible ellipsoidal
% possible ellipsoidal anomalous
% possible spherical
% possible spherical anomalous
Redundancy anomalous
% possible all
4.376-34.299
35.43
0.0391
94.16
121027
121027
3416
3416
0.999
-0.396
0.0065
0.0396
0.779
100
100
100
100
100
18.97
100
3.459-4.376
38.12
0.048
80.41
131526
131526
3450
3450
0.999
-0.464
0.0078
0.0487
0.74
100
100
100
100
100
19.81
100
3.015-3.459
41.19
0.0834
51.87
141112
141112
3426
3426
0.998
-0.402
0.0132
0.0844
0.754
100
100
100
100
100
21.3
100
2.733-3.015
36.96
0.1586
28.5
126102
126102
3412
3412
0.997
-0.125
0.0263
0.1608
0.757
100
100
100
100
100
19.05
100
2.537-2.733
39.44
0.3058
16.6
135679
135679
3440
3440
0.992
-0.049
0.0492
0.3098
0.759
100
100
100
100
100
20.26
100
2.385-2.537
41.19
0.5461
9.47
141174
141174
3427
3427
0.98
-0.022
0.0859
0.5529
0.74
100
100
100
100
100
21.11
100
2.264-2.385
41.84
0.83
6.17
143332
143332
3426
3426
0.953
-0.061
0.1295
0.8401
0.726
100
100
100
100
100
21.42
100
2.165-2.264
41.93
1.238
4.16
143689
143689
3427
3427
0.901
-0.057
0.1931
1.2531
0.73
100
100
100
100
100
21.45
100
2.08-2.165
42.5
1.9634
2.53
145978
145978
3435
3435
0.802
-0.007
0.3041
1.987
0.728
100
100
100
100
100
21.71
100
1.984-2.08
39.58
3.3256
1.45
135367
135367
3420
3420
0.588
-0.01
0.5339
3.3684
0.767
74.5
74.1
74.5
74.1
74.5
20.15
74.1
-
解析
ソフトウェア
名称
バージョン
分類
autoPROC
1.1.720220608
dataprocessing
XDS
Jan10, 2022
データ削減
Aimless
0.7.9
データスケーリング
STARANISO
2.3.87
データスケーリング
REFMAC
5.8.0352
精密化
REFMAC
5.8.0352
位相決定
精密化
構造決定の手法: フーリエ合成 / 解像度: 1.984→34.299 Å / Cor.coef. Fo:Fc: 0.968 / Cor.coef. Fo:Fc free: 0.956 / SU B: 7.143 / SU ML: 0.099 / 交差検証法: FREE R-VALUE / ESU R: 0.141 / ESU R Free: 0.131 / 詳細: Hydrogens have been added in their riding positions
Rfactor
反射数
%反射
Rfree
0.2156
1439
4.199 %
Rwork
0.1859
32835
-
all
0.187
-
-
obs
-
34274
96.538 %
溶媒の処理
イオンプローブ半径: 0.8 Å / 減衰半径: 0.8 Å / VDWプローブ半径: 1.2 Å / 溶媒モデル: MASK BULK SOLVENT
原子変位パラメータ
Biso mean: 57.55 Å2
Baniso -1
Baniso -2
Baniso -3
1-
0 Å2
0 Å2
0 Å2
2-
-
0 Å2
0 Å2
3-
-
-
0 Å2
精密化ステップ
サイクル: LAST / 解像度: 1.984→34.299 Å
タンパク質
核酸
リガンド
溶媒
全体
原子数
2606
0
38
217
2861
拘束条件
Refine-ID
タイプ
Dev ideal
Dev ideal target
数
X-RAY DIFFRACTION
r_bond_refined_d
0.005
0.012
2771
X-RAY DIFFRACTION
r_bond_other_d
0.002
0.016
2593
X-RAY DIFFRACTION
r_ext_dist_refined_b
0
0.01
3760
X-RAY DIFFRACTION
r_angle_refined_deg
1.315
1.672
3766
X-RAY DIFFRACTION
r_angle_other_deg
0.474
1.578
6049
X-RAY DIFFRACTION
r_dihedral_angle_1_deg
6.566
5
337
X-RAY DIFFRACTION
r_dihedral_angle_2_deg
3.941
5
18
X-RAY DIFFRACTION
r_dihedral_angle_3_deg
11.722
10
501
X-RAY DIFFRACTION
r_dihedral_angle_6_deg
13.781
10
119
X-RAY DIFFRACTION
r_chiral_restr
0.062
0.2
413
X-RAY DIFFRACTION
r_gen_planes_refined
0.006
0.02
3255
X-RAY DIFFRACTION
r_gen_planes_other
0.001
0.02
533
X-RAY DIFFRACTION
r_nbd_refined
0.209
0.2
493
X-RAY DIFFRACTION
r_symmetry_nbd_other
0.18
0.2
2496
X-RAY DIFFRACTION
r_nbtor_refined
0.168
0.2
1340
X-RAY DIFFRACTION
r_symmetry_nbtor_other
0.072
0.2
1384
X-RAY DIFFRACTION
r_xyhbond_nbd_refined
0.143
0.2
141
X-RAY DIFFRACTION
r_symmetry_xyhbond_nbd_other
0.003
0.2
1
X-RAY DIFFRACTION
r_symmetry_nbd_refined
0.215
0.2
22
X-RAY DIFFRACTION
r_nbd_other
0.149
0.2
89
X-RAY DIFFRACTION
r_symmetry_xyhbond_nbd_refined
0.187
0.2
21
X-RAY DIFFRACTION
r_mcbond_it
2.177
5.187
1331
X-RAY DIFFRACTION
r_mcbond_other
2.176
5.187
1331
X-RAY DIFFRACTION
r_mcangle_it
3.712
7.759
1673
X-RAY DIFFRACTION
r_mcangle_other
3.711
7.762
1674
X-RAY DIFFRACTION
r_scbond_it
2.029
5.4
1440
X-RAY DIFFRACTION
r_scbond_other
2.028
5.399
1441
X-RAY DIFFRACTION
r_scangle_it
3.458
7.983
2093
X-RAY DIFFRACTION
r_scangle_other
3.457
7.982
2094
X-RAY DIFFRACTION
r_lrange_it
7.078
98.965
5599
X-RAY DIFFRACTION
r_lrange_other
7.063
98.352
5559
LS精密化 シェル
解像度 (Å)
Rfactor Rfree
Num. reflection Rfree
Rfactor Rwork
Num. reflection Rwork
Refine-ID
% reflection obs (%)
1.984-2.036
0.254
66
0.331
1384
X-RAY DIFFRACTION
55.7692
2.036-2.091
0.288
106
0.287
2360
X-RAY DIFFRACTION
97.2781
2.091-2.152
0.248
109
0.242
2371
X-RAY DIFFRACTION
100
2.152-2.218
0.274
101
0.223
2275
X-RAY DIFFRACTION
100
2.218-2.29
0.264
93
0.22
2234
X-RAY DIFFRACTION
100
2.29-2.37
0.233
83
0.211
2155
X-RAY DIFFRACTION
100
2.37-2.459
0.256
99
0.204
2069
X-RAY DIFFRACTION
100
2.459-2.559
0.199
87
0.202
2005
X-RAY DIFFRACTION
100
2.559-2.672
0.231
73
0.196
1936
X-RAY DIFFRACTION
100
2.672-2.802
0.195
81
0.201
1831
X-RAY DIFFRACTION
100
2.802-2.952
0.231
64
0.185
1766
X-RAY DIFFRACTION
100
2.952-3.13
0.244
78
0.189
1665
X-RAY DIFFRACTION
100
3.13-3.344
0.194
68
0.183
1552
X-RAY DIFFRACTION
100
3.344-3.609
0.214
70
0.185
1458
X-RAY DIFFRACTION
100
3.609-3.949
0.206
61
0.174
1349
X-RAY DIFFRACTION
100
3.949-4.408
0.204
55
0.155
1221
X-RAY DIFFRACTION
100
4.408-5.076
0.17
53
0.151
1081
X-RAY DIFFRACTION
100
5.076-6.183
0.257
47
0.204
929
X-RAY DIFFRACTION
100
6.183-8.605
0.22
26
0.187
747
X-RAY DIFFRACTION
100
8.605-34.299
0.18
19
0.173
447
X-RAY DIFFRACTION
100
精密化 TLS
手法: refined / Origin x: 24.8463 Å / Origin y: 7.3736 Å / Origin z: 58.2865 Å
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) X線回折 /
X線回折 /  データ登録者
データ登録者 引用
引用 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード
 PDBj
PDBj











 集合体
集合体

 Spodoptera frugiperda (ツマジロクサヨトウ)
Spodoptera frugiperda (ツマジロクサヨトウ)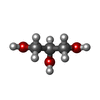
 分子量: 92.094 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C3H8O3
分子量: 92.094 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C3H8O3
 分子量: 35.453 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Cl
分子量: 35.453 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Cl 分子量: 18.015 Da / 分子数: 217 / 由来タイプ: 天然 / 式: H2O
分子量: 18.015 Da / 分子数: 217 / 由来タイプ: 天然 / 式: H2O 試料調製
試料調製 / ビームライン: MASSIF-1 / 波長: 0.965459 Å
/ ビームライン: MASSIF-1 / 波長: 0.965459 Å 解析
解析