 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8s1p | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | YlmH bound to PtRNA-50S | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Components Components |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Keywords Keywords | RIBOSOME / LSU / 50S / RQC / YlmH | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationRQC complex / positive regulation of rRNA processing / nucleoid / ribosomal large subunit binding / rescue of stalled cytosolic ribosome / rRNA processing / large ribosomal subunit / transferase activity / ribosomal small subunit biogenesis / 5S rRNA binding ...RQC complex / positive regulation of rRNA processing / nucleoid / ribosomal large subunit binding / rescue of stalled cytosolic ribosome / rRNA processing / large ribosomal subunit / transferase activity / ribosomal small subunit biogenesis / 5S rRNA binding / ribosomal large subunit assembly / small ribosomal subunit / large ribosomal subunit rRNA binding / cytosolic large ribosomal subunit / cytoplasmic translation / tRNA binding / negative regulation of translation / rRNA binding / structural constituent of ribosome / ribosome / translation / ribonucleoprotein complex / mRNA binding / DNA binding / RNA binding / metal ion binding / cytoplasm Similarity search - Function | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Biological species |  | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 1.96 Å | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Authors Authors | Paternoga, H. / Dimitrova-Paternoga, L. / Wilson, D.N. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Funding support |  Germany, 1items Germany, 1items
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Citation Citation | Journal: Nucleic Acids Res / Year: 2024 Title: A role for the S4-domain containing protein YlmH in ribosome-associated quality control in Bacillus subtilis. Authors: Hiraku Takada / Helge Paternoga / Keigo Fujiwara / Jose A Nakamoto / Esther N Park / Lyudmila Dimitrova-Paternoga / Bertrand Beckert / Merilin Saarma / Tanel Tenson / Allen R Buskirk / Gemma ...Authors: Hiraku Takada / Helge Paternoga / Keigo Fujiwara / Jose A Nakamoto / Esther N Park / Lyudmila Dimitrova-Paternoga / Bertrand Beckert / Merilin Saarma / Tanel Tenson / Allen R Buskirk / Gemma C Atkinson / Shinobu Chiba / Daniel N Wilson / Vasili Hauryliuk /      Abstract: Ribosomes trapped on mRNAs during protein synthesis need to be rescued for the cell to survive. The most ubiquitous bacterial ribosome rescue pathway is trans-translation mediated by tmRNA and SmpB. ...Ribosomes trapped on mRNAs during protein synthesis need to be rescued for the cell to survive. The most ubiquitous bacterial ribosome rescue pathway is trans-translation mediated by tmRNA and SmpB. Genetic inactivation of trans-translation can be lethal, unless ribosomes are rescued by ArfA or ArfB alternative rescue factors or the ribosome-associated quality control (RQC) system, which in Bacillus subtilis involves MutS2, RqcH, RqcP and Pth. Using transposon sequencing in a trans-translation-incompetent B. subtilis strain we identify a poorly characterized S4-domain-containing protein YlmH as a novel potential RQC factor. Cryo-EM structures reveal that YlmH binds peptidyl-tRNA-50S complexes in a position analogous to that of S4-domain-containing protein RqcP, and that, similarly to RqcP, YlmH can co-habit with RqcH. Consistently, we show that YlmH can assume the role of RqcP in RQC by facilitating the addition of poly-alanine tails to truncated nascent polypeptides. While in B. subtilis the function of YlmH is redundant with RqcP, our taxonomic analysis reveals that in multiple bacterial phyla RqcP is absent, while YlmH and RqcH are present, suggesting that in these species YlmH plays a central role in the RQC. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8s1p.cif.gz | 2.4 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8s1p.ent.gz | 1.8 MB | Display | PDB format |
| PDBx/mmJSON format | 8s1p.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/s1/8s1pftp://data.pdbj.org/pub/pdb/validation_reports/s1/8s1p | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  19638MC  8s1uC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |

-Links
 PDBj
PDBj





























- Assembly
Assembly
| Deposited unit | 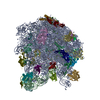
|
|---|---|
| 1 |
|
-Components
+50S ribosomal protein ... , 22 types, 22 molecules 346CDEFJKLMNOPRSUXY012
-RNA chain , 3 types, 3 molecules ABa
| #4: RNA chain | Mass: 949761.375 Da / Num. of mol.: 1 / Source method: isolated from a natural source Source: (natural) |
|---|---|
| #5: RNA chain | Mass: 36157.520 Da / Num. of mol.: 1 / Source method: isolated from a natural source Source: (natural) References: GenBank: 1150402534 |
| #29: RNA chain | Mass: 24491.547 Da / Num. of mol.: 1 / Source method: isolated from a natural source Source: (natural) References: GenBank: 1837880844 |
-Large ribosomal subunit protein ... , 5 types, 5 molecules GQTWZ
| #10: Protein | Mass: 19543.389 Da / Num. of mol.: 1 / Source method: isolated from a natural source Source: (natural) References: UniProt: P46898 |
|---|---|
| #18: Protein | Mass: 13669.189 Da / Num. of mol.: 1 / Source method: isolated from a natural source Source: (natural) References: UniProt: P55873 |
| #21: Protein | Mass: 10978.813 Da / Num. of mol.: 1 / Source method: isolated from a natural source Source: (natural) References: UniProt: P42924 |
| #23: Protein | Mass: 10391.855 Da / Num. of mol.: 1 / Source method: isolated from a natural source Source: (natural) References: UniProt: P05657 |
| #26: Protein | Mass: 6650.795 Da / Num. of mol.: 1 / Source method: isolated from a natural source Source: (natural) References: UniProt: P19947 |
-Protein/peptide / Protein , 2 types, 2 molecules bV
| #27: Protein/peptide | Mass: 358.434 Da / Num. of mol.: 1 / Source method: isolated from a natural source Source: (natural) |
|---|---|
| #28: Protein | Mass: 30823.840 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Gene: ylmH, BSU15410 / Production host: |
-Non-polymers , 5 types, 2981 molecules 

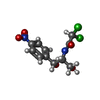


| #33: Chemical | ChemComp-ZN /  Mass: 65.409 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: Zn Mass: 65.409 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: Zn#34: Chemical | ChemComp-K /  Mass: 39.098 Da / Num. of mol.: 47 / Source method: obtained synthetically / Formula: K Mass: 39.098 Da / Num. of mol.: 47 / Source method: obtained synthetically / Formula: K#35: Chemical | ChemComp-CLM / |  Mass: 323.129 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C11H12Cl2N2O5 / Feature type: SUBJECT OF INVESTIGATION / Comment: antibiotic*YM Mass: 323.129 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C11H12Cl2N2O5 / Feature type: SUBJECT OF INVESTIGATION / Comment: antibiotic*YM#36: Chemical | ChemComp-MG /  Mass: 24.305 Da / Num. of mol.: 158 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 158 / Source method: obtained synthetically / Formula: Mg#37: Water | ChemComp-HOH / | Mass: 18.015 Da / Num. of mol.: 2771 / Source method: isolated from a natural source / Formula: H2O |
|---|
-Details
| Has ligand of interest | Y |
|---|---|
| Has protein modification | Y |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: 50S ribosomal subunit with stalled P-tRNA-nascent chain, bound by YlmH Type: RIBOSOME / Entity ID: #1-#32 / Source: NATURAL |
|---|---|
| Molecular weight | Experimental value: NO |
| Source (natural) | Organism: |
| Buffer solution | pH: 7.5 |
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Specimen support | Grid material: COPPER / Grid mesh size: 300 divisions/in. / Grid type: Quantifoil R3/3 |
| Vitrification | Cryogen name: ETHANE-PROPANE / Humidity: 100 % |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal defocus max: 900 nm / Nominal defocus min: 400 nm |
| Image recording | Electron dose: 40 e/Å2 / Film or detector model: FEI FALCON IV (4k x 4k) |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 2550984 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 1.96 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 66113 / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement | Resolution: 1.96→1.96 Å / Cor.coef. Fo:Fc: 0.884 / WRfactor Rwork: 0.286 / SU B: 4.668 / SU ML: 0.111 / Average fsc free: 0 / Average fsc overall: 0.837 / Average fsc work: 0.837 / ESU R: 0.111 Details: Hydrogens have been added in their riding positions
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Solvent model: NONE | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 57.13 Å2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
|