 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-8roo: Crystal structure of HLA B*18:01 in complex with YERMCNIL, an 8-m... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8roo | ||||||
|---|---|---|---|---|---|---|---|
| Title | Crystal structure of HLA B*18:01 in complex with YERMCNIL, an 8-mer epitope from Influenza A | ||||||
 Components Components |
| ||||||
 Keywords Keywords | IMMUNE SYSTEM / human leukocyte antigen / major histocompatibility complex / HLA-B*18:01 / Influenza A | ||||||
| Function / homology |  Function and homology information Function and homology informationnegative regulation of receptor binding / early endosome lumen / Nef mediated downregulation of MHC class I complex cell surface expression / DAP12 interactions / transferrin transport / cellular response to iron ion / Endosomal/Vacuolar pathway / Antigen Presentation: Folding, assembly and peptide loading of class I MHC / peptide antigen assembly with MHC class II protein complex / cellular response to iron(III) ion ...negative regulation of receptor binding / early endosome lumen / Nef mediated downregulation of MHC class I complex cell surface expression / DAP12 interactions / transferrin transport / cellular response to iron ion / Endosomal/Vacuolar pathway / Antigen Presentation: Folding, assembly and peptide loading of class I MHC / peptide antigen assembly with MHC class II protein complex / cellular response to iron(III) ion / MHC class II protein complex / negative regulation of forebrain neuron differentiation / antigen processing and presentation of exogenous protein antigen via MHC class Ib, TAP-dependent / ER to Golgi transport vesicle membrane / peptide antigen assembly with MHC class I protein complex / regulation of iron ion transport / regulation of erythrocyte differentiation / HFE-transferrin receptor complex / response to molecule of bacterial origin / MHC class I peptide loading complex / T cell mediated cytotoxicity / positive regulation of T cell cytokine production / antigen processing and presentation of endogenous peptide antigen via MHC class I / antigen processing and presentation of exogenous peptide antigen via MHC class II / positive regulation of immune response / MHC class I protein complex / positive regulation of T cell activation / peptide antigen binding / positive regulation of receptor-mediated endocytosis / negative regulation of neurogenesis / cellular response to nicotine / positive regulation of T cell mediated cytotoxicity / multicellular organismal-level iron ion homeostasis / specific granule lumen / phagocytic vesicle membrane / recycling endosome membrane / Interferon gamma signaling / Immunoregulatory interactions between a Lymphoid and a non-Lymphoid cell / negative regulation of epithelial cell proliferation / MHC class II protein complex binding / Modulation by Mtb of host immune system / late endosome membrane / sensory perception of smell / positive regulation of cellular senescence / tertiary granule lumen / DAP12 signaling / T cell differentiation in thymus / negative regulation of neuron projection development / ER-Phagosome pathway / protein refolding / early endosome membrane / protein homotetramerization / amyloid fibril formation / intracellular iron ion homeostasis / learning or memory / endoplasmic reticulum lumen / Amyloid fiber formation / Golgi membrane / lysosomal membrane / external side of plasma membrane / focal adhesion / Neutrophil degranulation / SARS-CoV-2 activates/modulates innate and adaptive immune responses / structural molecule activity / endoplasmic reticulum / Golgi apparatus / protein homodimerization activity / extracellular space / extracellular exosome / extracellular region / identical protein binding / membrane / plasma membrane / cytosol Similarity search - Function | ||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human)  Influenza A virus Influenza A virus | ||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 1.4 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 1.4 Å | ||||||
 Authors Authors | Murdolo, L.D. / Maddumage, J.C. / Gras, S. | ||||||
| Funding support |  Australia, 1items Australia, 1items
| ||||||
 Citation Citation | Journal: Clin Transl Immunology / Year: 2024 Title: Characterisation of novel influenza-derived HLA-B*18:01-restricted epitopes. Authors: Leong, S.L. / Murdolo, L. / Maddumage, J.C. / Koutsakos, M. / Kedzierska, K. / Purcell, A.W. / Gras, S. / Grant, E.J. #1: Journal: Acta Crystallogr D Struct Biol / Year: 2019 Title: Macromolecular structure determination using X-rays, neutrons and electrons: recent developments in Phenix. Authors: Dorothee Liebschner / Pavel V Afonine / Matthew L Baker / Gábor Bunkóczi / Vincent B Chen / Tristan I Croll / Bradley Hintze / Li Wei Hung / Swati Jain / Airlie J McCoy / Nigel W Moriarty ...Authors: Dorothee Liebschner / Pavel V Afonine / Matthew L Baker / Gábor Bunkóczi / Vincent B Chen / Tristan I Croll / Bradley Hintze / Li Wei Hung / Swati Jain / Airlie J McCoy / Nigel W Moriarty / Robert D Oeffner / Billy K Poon / Michael G Prisant / Randy J Read / Jane S Richardson / David C Richardson / Massimo D Sammito / Oleg V Sobolev / Duncan H Stockwell / Thomas C Terwilliger / Alexandre G Urzhumtsev / Lizbeth L Videau / Christopher J Williams / Paul D Adams /    Abstract: Diffraction (X-ray, neutron and electron) and electron cryo-microscopy are powerful methods to determine three-dimensional macromolecular structures, which are required to understand biological ...Diffraction (X-ray, neutron and electron) and electron cryo-microscopy are powerful methods to determine three-dimensional macromolecular structures, which are required to understand biological processes and to develop new therapeutics against diseases. The overall structure-solution workflow is similar for these techniques, but nuances exist because the properties of the reduced experimental data are different. Software tools for structure determination should therefore be tailored for each method. Phenix is a comprehensive software package for macromolecular structure determination that handles data from any of these techniques. Tasks performed with Phenix include data-quality assessment, map improvement, model building, the validation/rebuilding/refinement cycle and deposition. Each tool caters to the type of experimental data. The design of Phenix emphasizes the automation of procedures, where possible, to minimize repetitive and time-consuming manual tasks, while default parameters are chosen to encourage best practice. A graphical user interface provides access to many command-line features of Phenix and streamlines the transition between programs, project tracking and re-running of previous tasks. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8roo.cif.gz | 226.6 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8roo.ent.gz | 150.3 KB | Display | PDB format |
| PDBx/mmJSON format | 8roo.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Summary document | 8roo_validation.pdf.gz | 4.1 MB | Display | wwPDB validaton report |
|---|---|---|---|---|
| Full document | 8roo_full_validation.pdf.gz | 4.1 MB | Display | |
| Data in XML | 8roo_validation.xml.gz | 19.2 KB | Display | |
| Data in CIF | 8roo_validation.cif.gz | 28.4 KB | Display | |
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/ro/8rooftp://data.pdbj.org/pub/pdb/validation_reports/ro/8roo | HTTPS FTP |
-Related structure data

-Links
 PDBj
PDBj

- Assembly
Assembly
| Deposited unit | 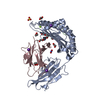
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||||||
| Unit cell |
|
-Components
-Protein , 2 types, 2 molecules AC
| #1: Protein | Mass: 31747.891 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: HLA-BProduction host:  References: UniProt: A0A167RQK8 |
|---|---|
| #3: Protein | Mass: 11879.356 Da / Num. of mol.: 1 / Fragment: UNP residues 21-119 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: B2M, CDABP0092, HDCMA22PProduction host: References: UniProt: P61769 |
-Protein/peptide , 1 types, 1 molecules B
| #2: Protein/peptide | Mass: 1042.254 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) Influenza A virus (A/X-31(H3N2)) |
|---|
-Non-polymers , 6 types, 314 molecules 



| #4: Chemical | ChemComp-EDO /  Mass: 62.068 Da / Num. of mol.: 9 / Source method: obtained synthetically / Formula: C2H6O2 / Feature type: SUBJECT OF INVESTIGATION Mass: 62.068 Da / Num. of mol.: 9 / Source method: obtained synthetically / Formula: C2H6O2 / Feature type: SUBJECT OF INVESTIGATION#5: Chemical | ChemComp-MG / |  Mass: 24.305 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Mg / Feature type: SUBJECT OF INVESTIGATION Mass: 24.305 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Mg / Feature type: SUBJECT OF INVESTIGATION#6: Chemical |  Mass: 39.098 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: K / Feature type: SUBJECT OF INVESTIGATION Mass: 39.098 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: K / Feature type: SUBJECT OF INVESTIGATION#7: Chemical | ChemComp-SO4 / |  Mass: 96.063 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: SO4 / Feature type: SUBJECT OF INVESTIGATION Mass: 96.063 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: SO4 / Feature type: SUBJECT OF INVESTIGATION#8: Chemical | ChemComp-NO3 / |  Mass: 62.005 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: NO3 / Feature type: SUBJECT OF INVESTIGATION Mass: 62.005 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: NO3 / Feature type: SUBJECT OF INVESTIGATION#9: Water | ChemComp-HOH / | Mass: 18.015 Da / Num. of mol.: 299 / Source method: isolated from a natural source / Formula: H2O |
|---|
-Details
| Has ligand of interest | Y |
|---|---|
| Has protein modification | Y |
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 2.58 Å3/Da / Density % sol: 52.35 % |
|---|---|
| Crystal grow | Temperature: 293 K / Method: vapor diffusion, sitting drop Details: 0.2 M Potassium Nitrate, 20% w/v PEG 3350 at pH 6.5 |
-Data collection
| Diffraction | Mean temperature: 100 K / Serial crystal experiment: N |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: Australian Synchrotron / Beamline: MX2 / Wavelength: 0.953732 Å |
| Detector | Type: DECTRIS EIGER X 16M / Detector: PIXEL / Date: Jul 19, 2023 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 0.953732 Å / Relative weight: 1 |
| Reflection | Resolution: 1.4→46.25 Å / Num. obs: 91186 / % possible obs: 99.6 % / Redundancy: 7.9 % / Biso Wilson estimate: 11.59 Å2 / CC1/2: 0.999 / Net I/σ(I): 15.6 |
| Reflection shell | Resolution: 1.4→1.42 Å / Mean I/σ(I) obs: 3.3 / Num. unique obs: 4394 / CC1/2: 0.639 |
- Processing
Processing
| Software |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT / Resolution: 1.4→40.74 Å / SU ML: 0.1326 / Cross valid method: FREE R-VALUE / σ(F): 1.35 / Phase error: 17.0392 Stereochemistry target values: GeoStd + Monomer Library + CDL v1.2
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.1 Å / Solvent model: FLAT BULK SOLVENT MODEL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 17.91 Å2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 1.4→40.74 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell |
|
