 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-8qvn: Cryo-EM structure of Cx26 from Gallus Gallus in bicarbonate buffer -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8qvn | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Cryo-EM structure of Cx26 from Gallus Gallus in bicarbonate buffer | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Components Components | Gap junction protein | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Keywords Keywords | MEMBRANE PROTEIN / Gap junction Large Pore Channel Carbon dioxide sensitive | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationGap junction assembly / gap junction-mediated intercellular transport / connexin complex / gap junction channel activity / sensory perception of sound / cell-cell signaling Similarity search - Function | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Biological species |  | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 2.07 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Authors Authors | Brotherton, D.H. / Cameron, A.D. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Funding support |  United Kingdom, 2items United Kingdom, 2items
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Citation Citation | Journal: J Physiol / Year: 2025 Title: Multiple carbamylation events are required for differential modulation of Cx26 hemichannels and gap junctions by CO. Authors: Sarbjit Nijjar / Deborah Brotherton / Jack Butler / Valentin-Mihai Dospinescu / Harry G Gannon / Victoria Linthwaite / Martin Cann / Alexander Cameron / Nicholas Dale / Abstract: CO directly modifies the gating of connexin26 (Cx26) gap junction channels and hemichannels. This gating depends upon Lys125, and the proposed mechanism involves carbamylation of Lys125 to allow ...CO directly modifies the gating of connexin26 (Cx26) gap junction channels and hemichannels. This gating depends upon Lys125, and the proposed mechanism involves carbamylation of Lys125 to allow formation of a salt bridge with Arg104 on the neighbouring subunit. We demonstrate via carbamate trapping and tandem mass spectrometry that five Lys residues within the cytoplasmic loop, including Lys125, are indeed carbamylated by CO. The cytoplasmic loop appears to provide a chemical microenvironment that facilitates carbamylation. Systematic mutation of these Lys residues to Arg shows that only carbamylation of Lys125 is essential for hemichannel opening. By contrast, carbamylation of Lys108 and Lys125 is essential for gap junction closure to CO. Chicken (Gallus gallus) Cx26 gap junction channels lack Lys108 and do not close to CO, as shown by both a dye transfer assay and a high-resolution cryogenic electron microscopy structure. The mutation Lys108Arg prevents CO-dependent gap junction channel closure in human Cx26. Our findings directly demonstrate carbamylation in connexins, provide further insight into the differential action of CO on Cx26 hemichannels and gap junction channels, and increase support for the role of the N-terminus in gating the Cx26 channel. KEY POINTS: Direct evidence of carbamylation of multiple lysine residues in the cytoplasmic loop of Cx26. Concentration-dependent carbamylation at lysines 108, 122 and 125. Only carbamylation of lysine 125 is essential for hemichannel opening to CO. Carbamylation of lysine 108 along with lysine 125 is essential for CO-dependent gap junction channel closure. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8qvn.cif.gz | 628.7 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8qvn.ent.gz | 419.7 KB | Display | PDB format |
| PDBx/mmJSON format | 8qvn.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/qv/8qvnftp://data.pdbj.org/pub/pdb/validation_reports/qv/8qvn | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  18670MC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |
-Links
 PDBj
PDBj- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
| #1: Protein | Mass: 26264.119 Da / Num. of mol.: 12 Source method: isolated from a genetically manipulated source Source: (gene. exp.)   Spodoptera frugiperda (fall armyworm) / References: UniProt: E1C2J9 Spodoptera frugiperda (fall armyworm) / References: UniProt: E1C2J9#2: Sugar | ChemComp-LMT / 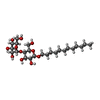  Type: D-saccharide / Mass: 510.615 Da / Num. of mol.: 24 / Source method: obtained synthetically / Formula: C24H46O11 / Comment: detergent*YM Type: D-saccharide / Mass: 510.615 Da / Num. of mol.: 24 / Source method: obtained synthetically / Formula: C24H46O11 / Comment: detergent*YM#3: Chemical | ChemComp-PTY /   Mass: 734.039 Da / Num. of mol.: 24 / Source method: obtained synthetically / Formula: C40H80NO8P / Comment: phospholipid*YM Mass: 734.039 Da / Num. of mol.: 24 / Source method: obtained synthetically / Formula: C40H80NO8P / Comment: phospholipid*YM#4: Water | ChemComp-HOH / |  Mass: 18.015 Da / Num. of mol.: 466 / Source method: isolated from a natural source / Formula: H2O Mass: 18.015 Da / Num. of mol.: 466 / Source method: isolated from a natural source / Formula: H2OHas ligand of interest | N | Has protein modification | Y | |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: Connexin 26 (Gallus Gallus) in 90mmHg PCO2, pH7.4 / Type: COMPLEX / Entity ID: #1 / Source: RECOMBINANT | |||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Experimental value: NO | |||||||||||||||||||||||||||||||||||||||||||||
| Source (natural) | Organism: | |||||||||||||||||||||||||||||||||||||||||||||
| Source (recombinant) | Organism: Spodoptera frugiperda (fall armyworm) | |||||||||||||||||||||||||||||||||||||||||||||
| Buffer solution | pH: 7.4 Details: The buffer, except DDM and DTT, was prepared fresh from 10x stock on day of used. The basal buffer was filtered and degassed and DDM and DTT added. The buffer was pH corrected at point of ...Details: The buffer, except DDM and DTT, was prepared fresh from 10x stock on day of used. The basal buffer was filtered and degassed and DDM and DTT added. The buffer was pH corrected at point of use to pH 7.4 using 15% CO2 in 85% N2. | |||||||||||||||||||||||||||||||||||||||||||||
| Buffer component |
| |||||||||||||||||||||||||||||||||||||||||||||
| Specimen | Conc.: 3 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | |||||||||||||||||||||||||||||||||||||||||||||
| Vitrification | Instrument: LEICA EM GP / Cryogen name: ETHANE-PROPANE Details: 3 microlitres protein applied to grid, blot time 7 seconds, in 15% CO2/85%N2 atmosphere |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 105000 X / Nominal defocus max: 2000 nm / Nominal defocus min: 800 nm / Cs: 2.7 mm |
| Specimen holder | Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Average exposure time: 3 sec. / Electron dose: 41 e/Å2 / Film or detector model: GATAN K3 (6k x 4k) / Num. of real images: 8713 |
| EM imaging optics | Energyfilter name: GIF Bioquantum |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 2041681 | ||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: D6 (2x6 fold dihedral) | ||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 2.07 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 161947 / Num. of class averages: 2 / Symmetry type: POINT | ||||||||||||||||||||||||||||||
| Atomic model building | Space: REAL | ||||||||||||||||||||||||||||||
| Atomic model building | PDB-ID: 7QEQ Accession code: 7QEQ / Source name: PDB / Type: experimental model | ||||||||||||||||||||||||||||||
| Refinement | Cross valid method: NONE Stereochemistry target values: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 84.6 Å2 | ||||||||||||||||||||||||||||||
| Refine LS restraints |
|

