 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-8jav: Structure of CRL2APPBP2 bound with the C-degron of MRPL28 (tetramer) -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8jav | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Structure of CRL2APPBP2 bound with the C-degron of MRPL28 (tetramer) | |||||||||||||||||||||||||||||||||
 Components Components |
| |||||||||||||||||||||||||||||||||
 Keywords Keywords | PROTEIN BINDING / E3 Ubiquitination ligase | |||||||||||||||||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationnegative regulation of beige fat cell differentiation / cullin-RING-type E3 NEDD8 transferase / NEDD8 transferase activity / cullin-RING ubiquitin ligase complex / Cul7-RING ubiquitin ligase complex / cellular response to chemical stress / target-directed miRNA degradation / Loss of Function of FBXW7 in Cancer and NOTCH1 Signaling / elongin complex / positive regulation of protein autoubiquitination ...negative regulation of beige fat cell differentiation / cullin-RING-type E3 NEDD8 transferase / NEDD8 transferase activity / cullin-RING ubiquitin ligase complex / Cul7-RING ubiquitin ligase complex / cellular response to chemical stress / target-directed miRNA degradation / Loss of Function of FBXW7 in Cancer and NOTCH1 Signaling / elongin complex / positive regulation of protein autoubiquitination / RNA polymerase II transcription initiation surveillance / protein neddylation / NEDD8 ligase activity / microtubule associated complex / protein K27-linked ubiquitination / microtubule motor activity / negative regulation of response to oxidative stress / VCB complex / Cul5-RING ubiquitin ligase complex / ubiquitin-ubiquitin ligase activity / ubiquitin-dependent protein catabolic process via the C-end degron rule pathway / SCF ubiquitin ligase complex / Cul2-RING ubiquitin ligase complex / Cul3-RING ubiquitin ligase complex / negative regulation of type I interferon production / SCF-dependent proteasomal ubiquitin-dependent protein catabolic process / Prolactin receptor signaling / Cul4A-RING E3 ubiquitin ligase complex / negative regulation of mitophagy / Cul4-RING E3 ubiquitin ligase complex / Cul4B-RING E3 ubiquitin ligase complex / ubiquitin ligase complex scaffold activity / Pausing and recovery of Tat-mediated HIV elongation / Tat-mediated HIV elongation arrest and recovery / HIV elongation arrest and recovery / Pausing and recovery of HIV elongation / intracellular transport / cullin family protein binding / protein monoubiquitination / Tat-mediated elongation of the HIV-1 transcript / Formation of HIV-1 elongation complex containing HIV-1 Tat / Formation of HIV elongation complex in the absence of HIV Tat / ubiquitin-like ligase-substrate adaptor activity / RNA Polymerase II Transcription Elongation / site of DNA damage / Formation of RNA Pol II elongation complex / signal transduction in response to DNA damage / Nuclear events stimulated by ALK signaling in cancer / protein K48-linked ubiquitination / RNA Polymerase II Pre-transcription Events / negative regulation of insulin receptor signaling pathway / regulation of cellular response to insulin stimulus / positive regulation of TORC1 signaling / transcription-coupled nucleotide-excision repair / post-translational protein modification / intrinsic apoptotic signaling pathway / cytoplasmic vesicle membrane / T cell activation / transcription corepressor binding / Regulation of BACH1 activity / negative regulation of canonical NF-kappaB signal transduction / TP53 Regulates Transcription of DNA Repair Genes / cellular response to amino acid stimulus / transcription initiation at RNA polymerase II promoter / transcription elongation by RNA polymerase II / negative regulation of canonical Wnt signaling pathway / G1/S transition of mitotic cell cycle / Degradation of DVL / Degradation of CRY and PER proteins / intracellular protein transport / Degradation of GLI1 by the proteasome / Recognition of DNA damage by PCNA-containing replication complex / GSK3B and BTRC:CUL1-mediated-degradation of NFE2L2 / Negative regulation of NOTCH4 signaling / RING-type E3 ubiquitin transferase / Hedgehog 'on' state / Inactivation of CSF3 (G-CSF) signaling / Vif-mediated degradation of APOBEC3G / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / Degradation of GLI2 by the proteasome / GLI3 is processed to GLI3R by the proteasome / Ubiquitin-Mediated Degradation of Phosphorylated Cdc25A / Evasion by RSV of host interferon responses / NOTCH1 Intracellular Domain Regulates Transcription / Degradation of beta-catenin by the destruction complex / DNA Damage Recognition in GG-NER / Oxygen-dependent proline hydroxylation of Hypoxia-inducible Factor Alpha / Constitutive Signaling by NOTCH1 PEST Domain Mutants / Constitutive Signaling by NOTCH1 HD+PEST Domain Mutants / Dual Incision in GG-NER / Transcription-Coupled Nucleotide Excision Repair (TC-NER) / Formation of TC-NER Pre-Incision Complex / Regulation of expression of SLITs and ROBOs / Formation of Incision Complex in GG-NER / Interleukin-1 signaling / Orc1 removal from chromatin / protein polyubiquitination / ubiquitin-protein transferase activity / Dual incision in TC-NER / positive regulation of protein catabolic process Similarity search - Function | |||||||||||||||||||||||||||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | |||||||||||||||||||||||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.44 Å | |||||||||||||||||||||||||||||||||
 Authors Authors | Zhao, S. / Zhang, K. / Xu, C. | |||||||||||||||||||||||||||||||||
| Funding support |  China, 1items China, 1items
| |||||||||||||||||||||||||||||||||
 Citation Citation | Journal: Proc Natl Acad Sci U S A / Year: 2023 Title: Molecular basis for C-degron recognition by CRL2 ubiquitin ligase. Authors: Shidong Zhao / Diana Olmayev-Yaakobov / Wenwen Ru / Shanshan Li / Xinyan Chen / Jiahai Zhang / Xuebiao Yao / Itay Koren / Kaiming Zhang / Chao Xu /  Abstract: E3 ubiquitin ligases determine the specificity of eukaryotic protein degradation by selective binding to destabilizing protein motifs, termed degrons, in substrates for ubiquitin-mediated proteolysis. ...E3 ubiquitin ligases determine the specificity of eukaryotic protein degradation by selective binding to destabilizing protein motifs, termed degrons, in substrates for ubiquitin-mediated proteolysis. The exposed C-terminal residues of proteins can act as C-degrons that are recognized by distinct substrate receptors (SRs) as part of dedicated cullin-RING E3 ubiquitin ligase (CRL) complexes. APPBP2, an SR of Cullin 2-RING ligase (CRL2), has been shown to recognize R-x-x-G/C-degron; however, the molecular mechanism of recognition remains elusive. By solving several cryogenic electron microscopy structures of active CRL2 bound with different R-x-x-G/C-degrons, we unveiled the molecular mechanisms underlying the assembly of the CRL2 dimer and tetramer, as well as C-degron recognition. The structural study, complemented by binding experiments and cell-based assays, demonstrates that APPBP2 specifically recognizes the R-x-x-G/C-degron via a bipartite mechanism; arginine and glycine, which play critical roles in C-degron recognition, accommodate distinct pockets that are spaced by two residues. In addition, the binding pocket is deep enough to enable the interaction of APPBP2 with the motif placed at or up to three residues upstream of the C-end. Overall, our study not only provides structural insight into CRL2-mediated protein turnover but also serves as the basis for future structure-based chemical probe design. | |||||||||||||||||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8jav.cif.gz | 906.5 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8jav.ent.gz | 729.8 KB | Display | PDB format |
| PDBx/mmJSON format | 8jav.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/ja/8javftp://data.pdbj.org/pub/pdb/validation_reports/ja/8jav | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  36135MC  8jalC 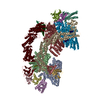 8jaqC  8jarC 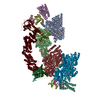 8jasC  8jauC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |




-Links
 PDBj
PDBj















- Assembly
Assembly
| Deposited unit | 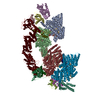
|
|---|---|
| 1 |
|
-Components
-Protein , 5 types, 18 molecules ABJKEILUCGMQDHNTRV
| #1: Protein | Mass: 66945.258 Da / Num. of mol.: 4 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: APPBP2, KIAA0228, PAT1 / Production host:  #2: Protein | Mass: 87068.836 Da / Num. of mol.: 4 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: CUL2 / Production host: #3: Protein | Mass: 13147.781 Da / Num. of mol.: 4 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: ELOB, TCEB2 / Production host: #4: Protein | Mass: 10843.420 Da / Num. of mol.: 4 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: ELOC, TCEB1 / Production host: #6: Protein | Mass: 12289.977 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: RBX1, RNF75, ROC1 / Production host: |
|---|
-Protein/peptide / Non-polymers , 2 types, 12 molecules SFOP
| #5: Protein/peptide | Mass: 1670.864 Da / Num. of mol.: 4 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Production host: #7: Chemical | ChemComp-ZN /  Mass: 65.409 Da / Num. of mol.: 8 / Source method: obtained synthetically / Formula: Zn / Feature type: SUBJECT OF INVESTIGATION Mass: 65.409 Da / Num. of mol.: 8 / Source method: obtained synthetically / Formula: Zn / Feature type: SUBJECT OF INVESTIGATION |
|---|
-Details
| Has ligand of interest | Y |
|---|---|
| Has protein modification | N |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: CRL2APPBP2 E3 liganse / Type: COMPLEX / Entity ID: #1-#6 / Source: MULTIPLE SOURCES |
|---|---|
| Molecular weight | Value: 400 kDa/nm / Experimental value: NO |
| Source (natural) | Organism: Homo sapiens (human) |
| Source (recombinant) | Organism: |
| Buffer solution | pH: 7.5 |
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: OTHER FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: OTHER |
| Electron lens | Mode: BRIGHT FIELD / Nominal defocus max: 2000 nm / Nominal defocus min: 800 nm |
| Image recording | Electron dose: 50 e/Å2 / Film or detector model: GATAN K3 (6k x 4k) |
- Processing
Processing
| Software | Name: PHENIX / Version: 1.20.1_4487: / Classification: refinement | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software | Name: PHENIX / Category: model refinement | ||||||||||||||||||||||||
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.44 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 113976 / Symmetry type: POINT | ||||||||||||||||||||||||
| Refine LS restraints |
|
