 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 8bhv | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
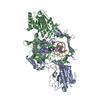
| タイトル | DNA-PK XLF mediated dimer bound to PAXX | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | DNA BINDING PROTEIN / DNA-PK / DNA-PKcs / Ku70 / Ku80 / PAXX / NHEJ | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報FHA domain binding / positive regulation of chromosome organization / positive regulation of ligase activity / DNA ligase IV complex / positive regulation of platelet formation / DNA ligase activity / Ku70:Ku80 complex / DN2 thymocyte differentiation / negative regulation of t-circle formation / DNA ligase (ATP) ...FHA domain binding / positive regulation of chromosome organization / positive regulation of ligase activity / DNA ligase IV complex / positive regulation of platelet formation / DNA ligase activity / Ku70:Ku80 complex / DN2 thymocyte differentiation / negative regulation of t-circle formation / DNA ligase (ATP) / T cell receptor V(D)J recombination / DNA end binding / pro-B cell differentiation / small-subunit processome assembly / positive regulation of lymphocyte differentiation / DNA ligase (ATP) activity / DNA-dependent protein kinase complex / DNA-dependent protein kinase-DNA ligase 4 complex / nonhomologous end joining complex / immunoglobulin V(D)J recombination / nucleotide-excision repair, DNA gap filling / immature B cell differentiation / DNA-dependent protein kinase activity / histone H2AXS139 kinase activity / single strand break repair / regulation of smooth muscle cell proliferation / positive regulation of double-strand break repair via nonhomologous end joining / cellular response to X-ray / V(D)J recombination / nuclear telomere cap complex / double-strand break repair via alternative nonhomologous end joining / double-strand break repair via classical nonhomologous end joining / protein localization to site of double-strand break / regulation of epithelial cell proliferation / telomere capping / isotype switching / Cytosolic sensors of pathogen-associated DNA / IRF3-mediated induction of type I IFN / positive regulation of neurogenesis / regulation of hematopoietic stem cell differentiation / regulation of telomere maintenance / recombinational repair / U3 snoRNA binding / protein localization to chromosome, telomeric region / DNA biosynthetic process / maturation of 5.8S rRNA / T cell lineage commitment / response to ionizing radiation / cellular response to lithium ion / cellular hyperosmotic salinity response / negative regulation of cGAS/STING signaling pathway / 2-LTR circle formation / telomeric DNA binding / B cell lineage commitment / hematopoietic stem cell proliferation / ligase activity / negative regulation of protein phosphorylation / positive regulation of protein kinase activity / site of DNA damage / 付加脱離酵素(リアーゼ); 炭素-酸素リアーゼ類; その他の炭素-酸素リアーゼ / T cell differentiation / peptidyl-threonine phosphorylation / somatic stem cell population maintenance / 5'-deoxyribose-5-phosphate lyase activity / response to X-ray / hematopoietic stem cell differentiation / ATP-dependent activity, acting on DNA / ectopic germ cell programmed cell death / chromosome organization / telomere maintenance via telomerase / somitogenesis / SUMOylation of DNA damage response and repair proteins / condensed chromosome / DNA polymerase binding / neurogenesis / mitotic G1 DNA damage checkpoint signaling / cellular response to ionizing radiation / telomere maintenance / DNA helicase activity / activation of innate immune response / cyclin binding / positive regulation of erythrocyte differentiation / positive regulation of translation / cellular response to leukemia inhibitory factor / B cell differentiation / central nervous system development / stem cell proliferation / response to gamma radiation / Nonhomologous End-Joining (NHEJ) / small-subunit processome / enzyme activator activity / cellular response to gamma radiation / regulation of circadian rhythm / protein destabilization / protein-DNA complex / base-excision repair / brain development / peptidyl-serine phosphorylation / protein modification process / 加水分解酵素; 酸無水物に作用; 酸無水物に作用・細胞または細胞小器官の運動に関与 類似検索 - 分子機能 | |||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 4.51 Å | |||||||||
 データ登録者 データ登録者 | Hardwick, S.W. / Chaplin, A.K. | |||||||||
| 資金援助 |  英国, 1件 英国, 1件
| |||||||||
 引用 引用 | ジャーナル: Sci Adv / 年: 2023 タイトル: PAXX binding to the NHEJ machinery explains functional redundancy with XLF. 著者: Murielle Seif-El-Dahan / Antonia Kefala-Stavridi / Philippe Frit / Steven W Hardwick / Dima Y Chirgadze / Taiana Maia De Oliviera / Jessica Andreani / Sébastien Britton / Nadia Barboule / ...著者: Murielle Seif-El-Dahan / Antonia Kefala-Stavridi / Philippe Frit / Steven W Hardwick / Dima Y Chirgadze / Taiana Maia De Oliviera / Jessica Andreani / Sébastien Britton / Nadia Barboule / Madeleine Bossaert / Arun Prasad Pandurangan / Katheryn Meek / Tom L Blundell / Virginie Ropars / Patrick Calsou / Jean-Baptiste Charbonnier / Amanda K Chaplin /   要旨: Nonhomologous end joining is a critical mechanism that repairs DNA double-strand breaks in human cells. In this work, we address the structural and functional role of the accessory protein PAXX ...Nonhomologous end joining is a critical mechanism that repairs DNA double-strand breaks in human cells. In this work, we address the structural and functional role of the accessory protein PAXX [paralog of x-ray repair cross-complementing protein 4 (XRCC4) and XRCC4-like factor (XLF)] in this mechanism. Here, we report high-resolution cryo-electron microscopy (cryo-EM) and x-ray crystallography structures of the PAXX C-terminal Ku-binding motif bound to Ku70/80 and cryo-EM structures of PAXX bound to two alternate DNA-dependent protein kinase (DNA-PK) end-bridging dimers, mediated by either Ku80 or XLF. We identify residues critical for the Ku70/PAXX interaction in vitro and in cells. We demonstrate that PAXX and XLF can bind simultaneously to the Ku heterodimer and act as structural bridges in alternate forms of DNA-PK dimers. Last, we show that engagement of both proteins provides a complementary advantage for DNA end synapsis and end joining in cells. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8bhv.cif.gz | 2 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8bhv.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 8bhv.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 8bhv_validation.pdf.gz | 1.7 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 8bhv_full_validation.pdf.gz | 2 MB | 表示 | |
| XML形式データ | 8bhv_validation.xml.gz | 316.2 KB | 表示 | |
| CIF形式データ | 8bhv_validation.cif.gz | 481.9 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/bh/8bhvftp://data.pdbj.org/pub/pdb/validation_reports/bh/8bhv | HTTPS FTP |
-関連構造データ









-リンク
 PDBj
PDBj












































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 5種, 12分子 AFKLNOMPQRci
| #1: タンパク質 | 分子量: 469673.219 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト)参照: UniProt: P78527, non-specific serine/threonine protein kinase #2: タンパク質 | 分子量: 38337.703 Da / 分子数: 4 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: XRCC4 / 発現宿主:  #3: タンパク質 | 分子量: 104124.953 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: LIG4 / 発現宿主: #4: タンパク質 | 分子量: 33372.234 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: NHEJ1, XLF / 発現宿主: #7: タンパク質 | 分子量: 21663.498 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PAXX, C9orf142, XLS / 発現宿主: |
|---|
-X-ray repair cross-complementing protein ... , 2種, 4分子 ahbj
| #5: タンパク質 | 分子量: 69945.039 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: XRCC6, G22P1発現宿主:   Spodoptera frugiperda (ツマジロクサヨトウ) Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: P12956, 加水分解酵素; 酸無水物に作用; 酸無水物に作用・細胞または細胞小器官の運動に関与, 付加脱離酵素(リアーゼ); 炭素- ...参照: UniProt: P12956, 加水分解酵素; 酸無水物に作用; 酸無水物に作用・細胞または細胞小器官の運動に関与, 付加脱離酵素(リアーゼ); 炭素-酸素リアーゼ類; その他の炭素-酸素リアーゼ #6: タンパク質 | 分子量: 82812.438 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: XRCC5, G22P2発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: P13010, 加水分解酵素; 酸無水物に作用; 酸無水物に作用・細胞または細胞小器官の運動に関与 |
|---|
-DNA鎖 , 4種, 4分子 DEIJ
| #8: DNA鎖 | 分子量: 8335.403 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) |
|---|---|
| #9: DNA鎖 | 分子量: 8619.629 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) |
| #10: DNA鎖 | 分子量: 7383.842 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) |
| #11: DNA鎖 | 分子量: 7362.797 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Homo sapiens (ヒト) |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: DNA-PK XLF mediated dimer bound to PAXX / タイプ: COMPLEX / Entity ID: all / 由来: MULTIPLE SOURCES |
|---|---|
| 分子量 | 値: 1.7 MDa / 実験値: NO |
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 緩衝液 | pH: 8 |
| 試料 | 濃度: 3 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 急速凍結 | 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 130000 X / 最大 デフォーカス(公称値): 2500 nm / 最小 デフォーカス(公称値): 800 nm / Cs: 2.7 mm / C2レンズ絞り径: 50 µm |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 電子線照射量: 48.03 e/Å2 フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) |
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 343564 | ||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 4.51 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 11019 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||
| 精密化 | 交差検証法: NONE 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 782.9 Å2 | ||||||||||||||||||||||||||||||
| 拘束条件 |
|
