 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 6ptn | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Structure of Ctf4 trimer in complex with two CMG helicases | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | REPLICATION / DNA replication / Cryo-EM / CMG-Ctf4 | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報establishment of sister chromatid cohesion / Unwinding of DNA / Cul8-RING ubiquitin ligase complex / MCM core complex / Assembly of the pre-replicative complex / Switching of origins to a post-replicative state / DNA strand elongation involved in mitotic DNA replication / MCM complex binding / GINS complex / nuclear DNA replication ...establishment of sister chromatid cohesion / Unwinding of DNA / Cul8-RING ubiquitin ligase complex / MCM core complex / Assembly of the pre-replicative complex / Switching of origins to a post-replicative state / DNA strand elongation involved in mitotic DNA replication / MCM complex binding / GINS complex / nuclear DNA replication / mitotic DNA replication preinitiation complex assembly / premeiotic DNA replication / pre-replicative complex assembly involved in nuclear cell cycle DNA replication / mitotic DNA replication / Activation of the pre-replicative complex / CMG complex / nuclear pre-replicative complex / Activation of ATR in response to replication stress / DNA replication preinitiation complex / MCM complex / replication fork protection complex / double-strand break repair via break-induced replication / mitotic DNA replication initiation / single-stranded DNA helicase activity / regulation of DNA-templated DNA replication initiation / DNA strand elongation involved in DNA replication / silent mating-type cassette heterochromatin formation / mitotic sister chromatid cohesion / nuclear chromosome / DNA unwinding involved in DNA replication / nuclear replication fork / DNA replication origin binding / DNA replication initiation / subtelomeric heterochromatin formation / DNA helicase activity / helicase activity / transcription elongation by RNA polymerase II / heterochromatin formation / DNA-templated DNA replication / nucleosome assembly / mitotic cell cycle / single-stranded DNA binding / DNA helicase / chromosome, telomeric region / DNA repair / DNA damage response / chromatin binding / ATP hydrolysis activity / nucleoplasm / ATP binding / identical protein binding / nucleus / metal ion binding / cytoplasm 類似検索 - 分子機能 | ||||||||||||
| 生物種 |  | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 5.8 Å | ||||||||||||
 データ登録者 データ登録者 | Yuan, Z. / Georgescu, R. / Bai, L. / Santos, R. / Donnell, M. / Li, H. | ||||||||||||
| 資金援助 |  米国, 3件 米国, 3件
| ||||||||||||
 引用 引用 | ジャーナル: Elife / 年: 2019 タイトル: Ctf4 organizes sister replisomes and Pol α into a replication factory. 著者: Zuanning Yuan / Roxana Georgescu / Ruda de Luna Almeida Santos / Daniel Zhang / Lin Bai / Nina Y Yao / Gongpu Zhao / Michael E O'Donnell / Huilin Li / 要旨: The current view is that eukaryotic replisomes are independent. Here we show that Ctf4 tightly dimerizes CMG helicase, with an extensive interface involving Psf2, Cdc45, and Sld5. Interestingly, Ctf4 ...The current view is that eukaryotic replisomes are independent. Here we show that Ctf4 tightly dimerizes CMG helicase, with an extensive interface involving Psf2, Cdc45, and Sld5. Interestingly, Ctf4 binds only one Pol α-primase. Thus, Ctf4 may have evolved as a trimer to organize two helicases and one Pol α-primase into a replication factory. In the 2CMG-Ctf4-1Pol α-primase factory model, the two CMGs nearly face each other, placing the two lagging strands toward the center and two leading strands out the sides. The single Pol α-primase is centrally located and may prime both sister replisomes. The Ctf4-coupled-sister replisome model is consistent with cellular microscopy studies revealing two sister forks of an origin remain attached and are pushed forward from a protein platform. The replication factory model may facilitate parental nucleosome transfer during replication. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 6ptn.cif.gz | 2.1 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb6ptn.ent.gz | 1.7 MB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 6ptn.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 6ptn_validation.pdf.gz | 1.4 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 6ptn_full_validation.pdf.gz | 1.4 MB | 表示 | |
| XML形式データ | 6ptn_validation.xml.gz | 254.4 KB | 表示 | |
| CIF形式データ | 6ptn_validation.cif.gz | 407.1 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/pt/6ptnftp://data.pdbj.org/pub/pdb/validation_reports/pt/6ptn | HTTPS FTP |
-関連構造データ










-リンク
 PDBj
PDBj
- 集合体
集合体
| 登録構造単位 | 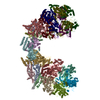
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 3種, 7分子 EFGhHl5
| #1: タンパク質 | 分子量: 104543.391 Da / 分子数: 3 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: CTF4, CHL15, POB1, YPR135W, P9659.7 / 発現宿主: #6: タンパク質 | 分子量: 74324.836 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: CDC45, SLD4, YLR103C, L8004.11 / 発現宿主: #10: タンパク質 | 分子量: 86505.734 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: MCM5, CDC46, YLR274W, L9328.1 / 発現宿主: |
|---|
-DNA replication complex GINS protein ... , 4種, 8分子 aAbBcCdD
| #2: タンパク質 | 分子量: 24230.576 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: PSF1, YDR013W, PZA208, YD8119.18 / 発現宿主: #3: タンパク質 | 分子量: 25096.807 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: PSF2, YJL072C, HRF213, J1086 / 発現宿主: #4: タンパク質 | 分子量: 21977.135 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: PSF3, YOL146W / 発現宿主: #5: タンパク質 | 分子量: 33983.617 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: SLD5, YDR489W / 発現宿主: |
|---|
-DNA replication licensing factor ... , 5種, 10分子 i2j3k4m6n7
| #7: タンパク質 | 分子量: 98911.539 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: MCM2, YBL023C, YBL0438 / 発現宿主: #8: タンパク質 | 分子量: 107653.508 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: MCM3, YEL032W, SYGP-ORF23 / 発現宿主: #9: タンパク質 | 分子量: 105138.375 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: MCM4, CDC54, HCD21, YPR019W, YP9531.13 / 発現宿主: #11: タンパク質 | 分子量: 113110.211 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: MCM6, YGL201C / 発現宿主: #12: タンパク質 | 分子量: 95049.875 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: MCM7, CDC47, YBR202W, YBR1441 / 発現宿主: |
|---|
-非ポリマー , 1種, 6分子 
| #13: 化合物 | ChemComp-ATP /  分子量: 507.181 Da / 分子数: 6 / 由来タイプ: 合成 / 式: C10H16N5O13P3 / タイプ: SUBJECT OF INVESTIGATION / コメント: ATP, エネルギー貯蔵分子*YM 分子量: 507.181 Da / 分子数: 6 / 由来タイプ: 合成 / 式: C10H16N5O13P3 / タイプ: SUBJECT OF INVESTIGATION / コメント: ATP, エネルギー貯蔵分子*YM |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|---|
| Has protein modification | Y |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: CMG-Ctf4 / タイプ: COMPLEX / Entity ID: #1-#12 / 由来: RECOMBINANT |
|---|---|
| 由来(天然) | 生物種: |
| 由来(組換発現) | 生物種: |
| 緩衝液 | pH: 7.5 |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 試料支持 | 詳細: unspecified |
| 急速凍結 | 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD |
| 撮影 | 電子線照射量: 50 e/Å2 フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) |
- 解析
解析
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION |
|---|---|
| 3次元再構成 | 解像度: 5.8 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 53853 / 対称性のタイプ: POINT |
| 原子モデル構築 | プロトコル: RIGID BODY FIT |
