 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-4jpo: 5A resolution structure of Proteasome Assembly Chaperone Hsm3 in ... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 4jpo | ||||||
|---|---|---|---|---|---|---|---|
| Title | 5A resolution structure of Proteasome Assembly Chaperone Hsm3 in complex with a C-terminal fragment of Rpt1 | ||||||
 Components Components |
| ||||||
 Keywords Keywords | CHAPERONE/HYDROLASE / Hsm3 / Chaperone / Proteasome / Protein Complex / CHAPERONE-HYDROLASE complex | ||||||
| Function / homology |  Function and homology information Function and homology informationproteasome regulatory particle assembly / proteasome-activating activity / proteasome regulatory particle, base subcomplex / Proteasome assembly / Cross-presentation of soluble exogenous antigens (endosomes) / TNFR2 non-canonical NF-kB pathway / Ubiquitin-Mediated Degradation of Phosphorylated Cdc25A / Regulation of PTEN stability and activity / CDK-mediated phosphorylation and removal of Cdc6 / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis ...proteasome regulatory particle assembly / proteasome-activating activity / proteasome regulatory particle, base subcomplex / Proteasome assembly / Cross-presentation of soluble exogenous antigens (endosomes) / TNFR2 non-canonical NF-kB pathway / Ubiquitin-Mediated Degradation of Phosphorylated Cdc25A / Regulation of PTEN stability and activity / CDK-mediated phosphorylation and removal of Cdc6 / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / KEAP1-NFE2L2 pathway / Neddylation / Orc1 removal from chromatin / MAPK6/MAPK4 signaling / Antigen processing: Ubiquitination & Proteasome degradation / positive regulation of RNA polymerase II transcription preinitiation complex assembly / Ub-specific processing proteases / mismatch repair / Neutrophil degranulation / protein folding chaperone / proteasome complex / positive regulation of protein catabolic process / ubiquitin-dependent protein catabolic process / proteasome-mediated ubiquitin-dependent protein catabolic process / ubiquitin protein ligase binding / ATP hydrolysis activity / ATP binding / nucleus / cytosol / cytoplasm Similarity search - Function | ||||||
| Biological species |  | ||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / molecular replacement / Resolution: 5 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / molecular replacement / Resolution: 5 Å | ||||||
 Authors Authors | Lovell, S. / Battaile, K.P. / Singh, R. / Roelofs, J. | ||||||
 Citation Citation | Journal: Nature / Year: 2013 Title: Reconfiguration of the proteasome during chaperone-mediated assembly. Authors: Soyeon Park / Xueming Li / Ho Min Kim / Chingakham Ranjit Singh / Geng Tian / Martin A Hoyt / Scott Lovell / Kevin P Battaile / Michal Zolkiewski / Philip Coffino / Jeroen Roelofs / Yifan ...Authors: Soyeon Park / Xueming Li / Ho Min Kim / Chingakham Ranjit Singh / Geng Tian / Martin A Hoyt / Scott Lovell / Kevin P Battaile / Michal Zolkiewski / Philip Coffino / Jeroen Roelofs / Yifan Cheng / Daniel Finley /  Abstract: The proteasomal ATPase ring, comprising Rpt1-Rpt6, associates with the heptameric α-ring of the proteasome core particle (CP) in the mature proteasome, with the Rpt carboxy-terminal tails inserting ...The proteasomal ATPase ring, comprising Rpt1-Rpt6, associates with the heptameric α-ring of the proteasome core particle (CP) in the mature proteasome, with the Rpt carboxy-terminal tails inserting into pockets of the α-ring. Rpt ring assembly is mediated by four chaperones, each binding a distinct Rpt subunit. Here we report that the base subassembly of the Saccharomyces cerevisiae proteasome, which includes the Rpt ring, forms a high-affinity complex with the CP. This complex is subject to active dissociation by the chaperones Hsm3, Nas6 and Rpn14. Chaperone-mediated dissociation was abrogated by a non-hydrolysable ATP analogue, indicating that chaperone action is coupled to nucleotide hydrolysis by the Rpt ring. Unexpectedly, synthetic Rpt tail peptides bound α-pockets with poor specificity, except for Rpt6, which uniquely bound the α2/α3-pocket. Although the Rpt6 tail is not visualized within an α-pocket in mature proteasomes, it inserts into the α2/α3-pocket in the base-CP complex and is important for complex formation. Thus, the Rpt-CP interface is reconfigured when the lid complex joins the nascent proteasome to form the mature holoenzyme. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 4jpo.cif.gz | 191.4 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb4jpo.ent.gz | 147.8 KB | Display | PDB format |
| PDBx/mmJSON format | 4jpo.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Summary document | 4jpo_validation.pdf.gz | 433.2 KB | Display | wwPDB validaton report |
|---|---|---|---|---|
| Full document | 4jpo_full_validation.pdf.gz | 447 KB | Display | |
| Data in XML | 4jpo_validation.xml.gz | 21.4 KB | Display | |
| Data in CIF | 4jpo_validation.cif.gz | 32.7 KB | Display | |
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/jp/4jpoftp://data.pdbj.org/pub/pdb/validation_reports/jp/4jpo | HTTPS FTP |
-Related structure data
| Related structure data |  5593C  5611C  5612C  5613C 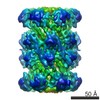 5614C  5615C  5616C  5617C  4fp7S S: Starting model for refinement C: citing same article ( |
|---|---|
| Similar structure data |








-Links
 PDBj
PDBj








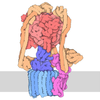





- Assembly
Assembly
| Deposited unit | 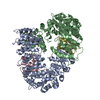
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||
| 2 | 
| ||||||||
| 3 | 
| ||||||||
| 4 | 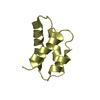
| ||||||||
| 5 | 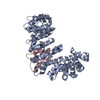
| ||||||||
| 6 | 
| ||||||||
| 7 |
| ||||||||
| 8 | 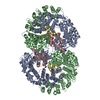
| ||||||||
| Unit cell |
|
-Components
| #1: Protein | Mass: 56685.719 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Strain: ATCC 204508 / S288c / Gene: HSM3, YBR272C, YBR1740 / Plasmid: pJR89 / Production host:  #2: Protein | Mass: 11637.442 Da / Num. of mol.: 2 / Fragment: C-terminal fragment, UNP residues 379-467 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Strain: ATCC 204508 / S288c / Gene: RPT1, CIM5, YTA3, YKL145W / Plasmid: pJR89 / Production host: |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal grow | Temperature: 293 K / Method: vapor diffusion / pH: 7 Details: 3M Sodium Formate, 100 mM Bis-Tris, 10mM Calcium Chloride, pH 7.0, vapor diffusion, temperature 293K |
|---|
-Data collection
| Diffraction | Mean temperature: 100 K |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: APS / Beamline: 17-ID / Wavelength: 1 Å |
| Detector | Type: DECTRIS PILATUS 6M / Detector: PIXEL / Date: Jan 1, 2011 |
| Radiation | Protocol: SINGLE WAVELENGTH / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 1 Å / Relative weight: 1 |
| Reflection | Resolution: 5→160.474 Å / Num. all: 16422 / Num. obs: 16422 / % possible obs: 99.9 % / Observed criterion σ(F): 0 / Observed criterion σ(I): -3 / Redundancy: 19.3 % / Rmerge(I) obs: 0.083 / Net I/σ(I): 20.1 |
| Reflection shell | Resolution: 5→5.59 Å / Redundancy: 19.2 % / Rmerge(I) obs: 0.971 / Mean I/σ(I) obs: 3.6 / Num. unique all: 4550 / % possible all: 100 |
-Phasing
| Phasing | Method: molecular replacement |
|---|
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: PDB ENTRY 4FP7 Resolution: 5→46.87 Å / Cor.coef. Fo:Fc: 0.9254 / Cor.coef. Fo:Fc free: 0.9109 / Occupancy max: 1 / Occupancy min: 1 / Cross valid method: THROUGHOUT / σ(F): 0 / Stereochemistry target values: Engh & Huber Details: B-factors were set to the value determined from the Wilson plot for refinement.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso max: 273 Å2 / Biso mean: 273 Å2 / Biso min: 273 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine analyze | Luzzati coordinate error obs: 2.055 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 5→46.87 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell | Resolution: 5→5.34 Å / Total num. of bins used: 8
|
