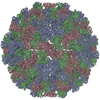
ジャーナル: J Mol Biol / 年: 1986 タイトル: Structure and assembly of turnip crinkle virus. I. X-ray crystallographic structure analysis at 3.2 A resolution. 要旨: The structure of turnip crinkle virus has been determined at 3.2 A resolution, using the electron density of tomato bushy stunt virus as a starting point for phase refinement by non-crystallographic ...The structure of turnip crinkle virus has been determined at 3.2 A resolution, using the electron density of tomato bushy stunt virus as a starting point for phase refinement by non-crystallographic symmetry. The structures are very closely related, especially in the subunit arm and S domain, where only small insertions and deletions and small co-ordinate shifts relate one chain to another. The P domains, although quite similar in fold, are oriented somewhat differently with respect to the S domains. Understanding of the structure of turnip crinkle virus has been important for analyzing its assembly, as described in an accompanying paper.
履歴
登録
2011年8月8日
登録サイト: PDBE / 処理サイト: PDBE
改定 1.0
2012年2月8日
Provider: repository / タイプ: Initial release
改定 1.1
2015年3月4日
Group: Database references
改定 1.2
2016年2月3日
Group: Derived calculations
改定 1.3
2017年6月28日
Group: Data collection / カテゴリ: diffrn_source Item: _diffrn_source.pdbx_wavelength_list / _diffrn_source.type
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報 TURNIP CRINKLE VIRUS (ウイルス)
TURNIP CRINKLE VIRUS (ウイルス) X線回折 /
X線回折 /  データ登録者
データ登録者 引用
引用 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード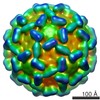




 PDBj
PDBj

 集合体
集合体



