contractile muscle fiber / regulation of double-strand break repair via nonhomologous end joining / Transcriptional regulation by the AP-2 (TFAP2) family of transcription factors / negative regulation of chromosome condensation / Barr body / : / B-WICH complex / pericentric heterochromatin formation / inner kinetochore / muscle cell differentiation ...contractile muscle fiber / regulation of double-strand break repair via nonhomologous end joining / Transcriptional regulation by the AP-2 (TFAP2) family of transcription factors / negative regulation of chromosome condensation / Barr body / : / B-WICH complex / pericentric heterochromatin formation / inner kinetochore / muscle cell differentiation / regulation of double-strand break repair / oocyte maturation / nucleosomal DNA binding / positive regulation of transcription by RNA polymerase III / nucleus organization / positive regulation of transcription by RNA polymerase I / spermatid development / negative regulation of tumor necrosis factor-mediated signaling pathway / single fertilization / subtelomeric heterochromatin formation / RNA polymerase II core promoter sequence-specific DNA binding / negative regulation of megakaryocyte differentiation / protein localization to CENP-A containing chromatin / Replacement of protamines by nucleosomes in the male pronucleus / CENP-A containing nucleosome / Packaging Of Telomere Ends / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / Deposition of new CENPA-containing nucleosomes at the centromere / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / embryo implantation / telomere organization / Inhibition of DNA recombination at telomere / RNA Polymerase I Promoter Opening / Meiotic synapsis / Assembly of the ORC complex at the origin of replication / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / SUMOylation of chromatin organization proteins / viral genome replication / DNA methylation / Condensation of Prophase Chromosomes / Chromatin modifications during the maternal to zygotic transition (MZT) / SIRT1 negatively regulates rRNA expression / HCMV Late Events / innate immune response in mucosa / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / PRC2 methylates histones and DNA / Regulation of endogenous retroelements by KRAB-ZFP proteins / Defective pyroptosis / HDACs deacetylate histones / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / Nonhomologous End-Joining (NHEJ) / lipopolysaccharide binding / RNA Polymerase I Promoter Escape / Transcriptional regulation by small RNAs / Formation of the beta-catenin:TCF transactivating complex / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / HDMs demethylate histones / G2/M DNA damage checkpoint / NoRC negatively regulates rRNA expression / DNA Damage/Telomere Stress Induced Senescence / B-WICH complex positively regulates rRNA expression / PKMTs methylate histone lysines / Meiotic recombination / Pre-NOTCH Transcription and Translation / Metalloprotease DUBs / multicellular organism growth / RMTs methylate histone arginines / Activation of anterior HOX genes in hindbrain development during early embryogenesis / male gonad development / Transcriptional regulation of granulopoiesis / HCMV Early Events / antimicrobial humoral immune response mediated by antimicrobial peptide / osteoblast differentiation / structural constituent of chromatin / UCH proteinases / antibacterial humoral response / nucleosome / heterochromatin formation / nucleosome assembly / E3 ubiquitin ligases ubiquitinate target proteins / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / RUNX1 regulates transcription of genes involved in differentiation of HSCs / HATs acetylate histones / Factors involved in megakaryocyte development and platelet production / chromatin organization / MLL4 and MLL3 complexes regulate expression of PPARG target genes in adipogenesis and hepatic steatosis / Processing of DNA double-strand break ends / Senescence-Associated Secretory Phenotype (SASP) / positive regulation of cell growth / Oxidative Stress Induced Senescence / histone binding / defense response to Gram-negative bacterium / killing of cells of another organism / Estrogen-dependent gene expression / transcription by RNA polymerase II / chromosome, telomeric region / cell population proliferation 類似検索 - 分子機能
Protein DEK / DEK C terminal domain / DEK, C-terminal / DEK C-terminal (DEK-C) domain profile. / SAP domain / Putative DNA-binding (bihelical) motif predicted to be involved in chromosomal organisation / SAP domain / : / Histone H2B signature. / Histone H2A conserved site ...Protein DEK / DEK C terminal domain / DEK, C-terminal / DEK C-terminal (DEK-C) domain profile. / SAP domain / Putative DNA-binding (bihelical) motif predicted to be involved in chromosomal organisation / SAP domain / : / Histone H2B signature. / Histone H2A conserved site / Histone H2A signature. / Histone H2B / Histone H2B / Histone H2A, C-terminal domain / C-terminus of histone H2A / Histone 2A / Histone H2A / TATA box binding protein associated factor / TATA box binding protein associated factor (TAF), histone-like fold domain / Histone H4, conserved site / Histone H4 signature. / Histone H4 / Histone H4 / CENP-T/Histone H4, histone fold / Centromere kinetochore component CENP-T histone fold / Histone H3 signature 1. / Histone H3 signature 2. / Histone H3 / Histone H3/CENP-A / Histone H2A/H2B/H3 / Core histone H2A/H2B/H3/H4 domain / Histone-fold 類似検索 - ドメイン・相同性
Histone H2A type 1-B/E / Histone H2B type 1-J / Protein DEK / Histone H4 / Histone H3.3 類似検索 - 構成要素
National Natural Science Foundation of China (NSFC)
32171206
中国
引用
ジャーナル: Nat Struct Mol Biol / 年: 2025 タイトル: DEK-nucleosome structure shows DEK modulates H3K27me3 and stem cell fate. 著者: Yunfan Shen / Yanhong Liu / Maochao Guo / Song Mao / Rui Chen / Mengran Wang / Zhengbo Li / Yue Li / Wan Chen / Fang Chen / Baixing Wu / Chongyuan Wang / Wei Chen / Huanhuan Cui / Kai Yuan / Hongda Huang / 要旨: DEK is a highly conserved chromatin-associated oncoprotein that has important roles in regulating chromatin dynamics and stem cell fate. Dysregulation of DEK is associated with stem cell dysfunction ...DEK is a highly conserved chromatin-associated oncoprotein that has important roles in regulating chromatin dynamics and stem cell fate. Dysregulation of DEK is associated with stem cell dysfunction and cancers, including acute myeloid leukemia. Despite its importance in chromatin regulation, the structural mechanisms underlying DEK's interaction with chromatin and its influence on gene regulation remain poorly understood. Here we combined cryogenic electron microscopy (cryo-EM), biochemical and cellular approaches to investigate the molecular mechanisms and functional importance of DEK's interaction with chromatin. Our cryo-EM structures reveal the structural basis of the DEK-nucleosome interaction. Biochemical and cellular results demonstrate that this interaction is crucial for DEK deposition onto chromatin. Furthermore, our results reveal that DEK safeguards mouse embryonic stem cells from acquiring primitive endoderm fates by modulating the repressive histone mark H3K27me3. Together, our study provides crucial molecular insights into the structure and function of DEK, establishing a framework for understanding its roles in chromatin biology and cell fate determination.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報
 マップデータ
マップデータ 試料
試料 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) データ登録者
データ登録者 中国, 1件
中国, 1件  引用
引用 構造の表示
構造の表示
 ダウンロードとリンク
ダウンロードとリンク emd_62757.png
emd_62757.png http://ftp.pdbj.org/pub/emdb/structures/EMD-62757
http://ftp.pdbj.org/pub/emdb/structures/EMD-62757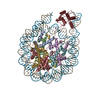

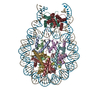









 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





































 試料の構成要素
試料の構成要素
 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN
