cytoplasmic U snRNP body / mRNA 3'-end processing by stem-loop binding and cleavage / co-transcriptional mRNA 3'-end processing, cleavage and polyadenylation pathway / 5'-3' RNA exonuclease activity / Processing of Intronless Pre-mRNAs / mRNA cleavage and polyadenylation specificity factor complex / U2 snRNP binding / U7 snRNA binding / histone pre-mRNA DCP binding / nuclear stress granule ...cytoplasmic U snRNP body / mRNA 3'-end processing by stem-loop binding and cleavage / co-transcriptional mRNA 3'-end processing, cleavage and polyadenylation pathway / 5'-3' RNA exonuclease activity / Processing of Intronless Pre-mRNAs / mRNA cleavage and polyadenylation specificity factor complex / U2 snRNP binding / U7 snRNA binding / histone pre-mRNA DCP binding / nuclear stress granule / U7 snRNP / regulation of chromatin organization / histone pre-mRNA 3'end processing complex / SLBP independent Processing of Histone Pre-mRNAs / SLBP Dependent Processing of Replication-Dependent Histone Pre-mRNAs / protein methylation / 7-methylguanosine cap hypermethylation / U12-type spliceosomal complex / U1 snRNP binding / 加水分解酵素; エステル加水分解酵素; 3'-リン酸モノエステル産生エンドリボヌクレアーゼ / methylosome / pICln-Sm protein complex / mRNA 3'-end processing / Transport of Mature mRNA Derived from an Intronless Transcript / small nuclear ribonucleoprotein complex / SMN-Sm protein complex / spliceosomal tri-snRNP complex / P granule / snRNP binding / commitment complex / U2-type precatalytic spliceosome / telomerase holoenzyme complex / mRNA 3'-end processing / telomerase RNA binding / U2-type catalytic step 2 spliceosome / U2-type prespliceosome assembly / U2-type spliceosomal complex / U1 snRNP / RNA Polymerase II Transcription Termination / U2 snRNP / U4 snRNP / U2-type prespliceosome / precatalytic spliceosome / mRNA Splicing - Minor Pathway / spliceosomal complex assembly / Processing of Capped Intron-Containing Pre-mRNA / U5 snRNP / bicellular tight junction / negative regulation of protein binding / spliceosomal snRNP assembly / positive regulation of G1/S transition of mitotic cell cycle / Cajal body / U4/U6 x U5 tri-snRNP complex / catalytic step 2 spliceosome / RNA endonuclease activity / mRNA Splicing - Major Pathway / RNA splicing / spliceosomal complex / mRNA splicing, via spliceosome / mRNA processing / snRNP Assembly / cytoskeleton / SARS-CoV-2 modulates host translation machinery / cell adhesion / postsynapse / nuclear body / ribonucleoprotein complex / glutamatergic synapse / enzyme binding / RNA binding / nucleoplasm / membrane / metal ion binding / nucleus / plasma membrane / cytoplasm / cytosol 類似検索 - 分子機能
Sm-like protein Lsm11, middle domain / U7 snRNA-associated Sm-like protein Lsm11 / : / Cleavage and polyadenylation specificity factor 2, C-terminal / CPSF2, metallo-hydrolase domain / Cleavage and polyadenylation factor 2 C-terminal / Cleavage and polyadenylation specificity factor subunit 2 / Pre-mRNA 3'-end-processing endonuclease polyadenylation factor C-term / Pre-mRNA 3'-end-processing endonuclease polyadenylation factor C-term / CPSF73-100_C ...Sm-like protein Lsm11, middle domain / U7 snRNA-associated Sm-like protein Lsm11 / : / Cleavage and polyadenylation specificity factor 2, C-terminal / CPSF2, metallo-hydrolase domain / Cleavage and polyadenylation factor 2 C-terminal / Cleavage and polyadenylation specificity factor subunit 2 / Pre-mRNA 3'-end-processing endonuclease polyadenylation factor C-term / Pre-mRNA 3'-end-processing endonuclease polyadenylation factor C-term / CPSF73-100_C / Symplekin/Pta1 / Symplekin C-terminal / Symplekin/Pta1, N-terminal / Symplekin/PTA1 N-terminal / Symplekin tight junction protein C terminal / Metallo-beta-lactamase superfamily domain / : / Beta-Casp domain / Beta-Casp domain / Beta-Casp domain / Zn-dependent metallo-hydrolase, RNA specificity domain / Zn-dependent metallo-hydrolase RNA specificity domain / Small ribonucleoprotein associated, SmB/SmN / Small nuclear ribonucleoprotein Sm D3 / Small nuclear ribonucleoprotein E / Small nuclear ribonucleoprotein G / Small nuclear ribonucleoprotein F / Sm-like protein Lsm7/SmG / Like-Sm (LSM) domain containing protein, LSm4/SmD1/SmD3 / Sm-like protein Lsm6/SmF / LSM domain / LSM domain, eukaryotic/archaea-type / snRNP Sm proteins / : / Sm domain profile. / LSM domain superfamily / Metallo-beta-lactamase superfamily / Metallo-beta-lactamase superfamily / Metallo-beta-lactamase / Ribonuclease Z/Hydroxyacylglutathione hydrolase-like / Armadillo-like helical / Armadillo-type fold 類似検索 - ドメイン・相同性
Small nuclear ribonucleoprotein-associated proteins B and B' / Small nuclear ribonucleoprotein E / Small nuclear ribonucleoprotein F / Small nuclear ribonucleoprotein G / Small nuclear ribonucleoprotein Sm D3 / U7 snRNA-associated Sm-like protein LSm11 / Symplekin / U7 snRNA-associated Sm-like protein LSm10 / Cleavage and polyadenylation specificity factor subunit 2 / Cleavage and polyadenylation specificity factor subunit 3 類似検索 - 構成要素
National Institutes of Health/National Institute of General Medical Sciences (NIH/NIGMS)
R35GM118093
米国
National Institutes of Health/National Institute of General Medical Sciences (NIH/NIGMS)
R01GM029832
米国
引用
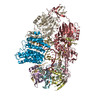
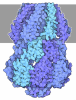
ジャーナル: Nucleic Acids Res / 年: 2026 タイトル: An N-terminal helix of Lsm11 stabilizes CPSF73 in U7 snRNP for histone pre-mRNA 3'-end processing. 著者: Anthony Desotell / William F Marzluff / Zbigniew Dominski / Liang Tong / 要旨: The U7 snRNP (small nuclear ribonucleoprotein) is responsible for the 3'-end processing of replication-dependent histone messenger RNA precursors (pre-mRNAs). A helix in the Lsm11 N-terminal ...The U7 snRNP (small nuclear ribonucleoprotein) is responsible for the 3'-end processing of replication-dependent histone messenger RNA precursors (pre-mRNAs). A helix in the Lsm11 N-terminal extension contacts the metallo-β-lactamase domain of the U7 snRNP endonuclease CPSF73. We mutated or deleted this helix and found that the mutant machineries had substantially reduced cleavage activity toward the pre-mRNA. Our cryo-electron microscopy (cryo-EM) studies indicated that the helix was important for helping to hold CPSF73 in its correct position for the cleavage reaction. We also reconstituted a wild-type U7 snRNP in complex with a methylated, noncleavable pre-mRNA. We observed that CPSF73 could achieve an open conformation independent of RNA binding to its active site. Finally, we found that a previously uninterpreted EM density for a small helix at the CPSF73-CPSF100 interface belonged to the C-terminal end of CstF77, copurified from insect cells and highly conserved among CstF77 homologs. This CstF77 binding site had a small effect on the cleavage activity of U7 snRNP. Overall, our studies have revealed the importance of the conserved helix in the Lsm11 N-terminal extension for U7 snRNP, provided structural evidence that CPSF73 can achieve an open, active conformation without RNA binding in its active site, and identified a previously unknown binding site for CstF77 in CPSF100.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報
 マップデータ
マップデータ 試料
試料 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト) /
Homo sapiens (ヒト) /  Trichoplusia ni (イラクサキンウワバ)
Trichoplusia ni (イラクサキンウワバ) データ登録者
データ登録者 米国, 2件
米国, 2件  引用
引用 構造の表示
構造の表示
 ダウンロードとリンク
ダウンロードとリンク emd_49402.png
emd_49402.png http://ftp.pdbj.org/pub/emdb/structures/EMD-49402
http://ftp.pdbj.org/pub/emdb/structures/EMD-49402














 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





































 試料の構成要素
試料の構成要素
 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN