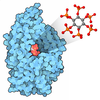
登録情報 データベース : EMDB / ID : EMD-48799タイトル PP2A-B55 Holoenzyme with B55i 複合体 : PP2A-B55 and B55i complexタンパク質・ペプチド : Serine/threonine-protein phosphatase 2A 65 kDa regulatory subunit A alpha isoformタンパク質・ペプチド : Serine/threonine-protein phosphatase 2A 55 kDa regulatory subunit B alpha isoformタンパク質・ペプチド : Serine/threonine-protein phosphatase 2A catalytic subunit alpha isoformタンパク質・ペプチド : B55i inhibitor peptide / / / / / 機能・相同性 分子機能 ドメイン・相同性 構成要素
/ / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / 生物種 Homo sapiens (ヒト)手法 / / 解像度 : 3.5 Å Shi S / Alderman C / Huang W / Zhao R 資金援助 Organization Grant number 国 National Institutes of Health/National Institute of General Medical Sciences (NIH/NIGMS) R35GM145289
ジャーナル : J Biol Chem / 年 : 2025タイトル : Cryo-EM structures reveal the PP2A-B55α and Eya3 interaction that can be disrupted by a peptide inhibitor.著者 : Shasha Shi / Xueni Li / Christopher Alderman / Lars Wick / Wei Huang / North Foulon / Lingdi Zhang / John Rossi / Wenxin Hu / Shouqing Cui / Hongjin Zheng / Derek J Taylor / Heide L Ford / Rui Zhao / 要旨 : We have previously shown that Eya3 recruits PP2A-B55α to dephosphorylate pT58 on Myc, increasing Myc stability and enhancing primary tumor growth of triple-negative breast cancer (TNBC). However, ... We have previously shown that Eya3 recruits PP2A-B55α to dephosphorylate pT58 on Myc, increasing Myc stability and enhancing primary tumor growth of triple-negative breast cancer (TNBC). However, the molecular details of how Eya3 recruits PP2A-B55α remain unclear. Here we determined the cryo-EM structures of PP2A-B55α bound with Eya3, with an inhibitory peptide B55i, and in its unbound state. These studies demonstrate that Eya3 binds B55α through an extended peptide in the NTD of Eya3. The Eya3 peptide, PP2A-B55α substrates, and protein/peptide inhibitors including B55i bind to a similar area on the B55α surface but the molecular details of the binding differ. We further demonstrated that the B55i peptide inhibits the B55α and Eya3 interaction in vitro. The B55i peptide expressed on a plasmid increases Myc pT58 and decreases Myc protein levels in TNBC cells, suggesting the potential of B55i or similar peptides as therapies for TNBC. 履歴 登録 2025年1月24日 - ヘッダ(付随情報) 公開 2025年6月18日 - マップ公開 2025年6月18日 - 更新 2025年6月18日 - 現状 2025年6月18日 処理サイト : RCSB / 状態 : 公開
すべて表示 表示を減らす
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報
 マップデータ
マップデータ 試料
試料 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) データ登録者
データ登録者 米国, 1件
米国, 1件  引用
引用 構造の表示
構造の表示
 ダウンロードとリンク
ダウンロードとリンク emd_48799.png
emd_48799.png http://ftp.pdbj.org/pub/emdb/structures/EMD-48799
http://ftp.pdbj.org/pub/emdb/structures/EMD-48799



















 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





































 試料の構成要素
試料の構成要素 解析
解析 電子顕微鏡法
電子顕微鏡法

