 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 |  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Structure of the LPD-3 complex | |||||||||
 マップデータ マップデータ | ||||||||||
 試料 試料 |
| |||||||||
 キーワード キーワード | Native / membrane protein complex / BLTP / LIPID TRANSPORT | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報 | |||||||||
| 生物種 |  | |||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 2.7 Å | |||||||||
 データ登録者 データ登録者 | Clark SA / Vanni S / Kang Y | |||||||||
| 資金援助 |  米国, 1件 米国, 1件
| |||||||||
 引用 引用 | ジャーナル: Nature / 年: 2025 タイトル: Structural basis of lipid transfer by a bridge-like lipid-transfer protein. 著者: Yunsik Kang / Katherine S Lehmann / Hannah Long / Amanda Jefferson / Maria Purice / Marc Freeman / Sarah Clark / 要旨: Bridge-like lipid-transport proteins (BLTPs) are an evolutionarily conserved family of proteins that localize to membrane-contact sites and are thought to mediate the bulk transfer of lipids from a ...Bridge-like lipid-transport proteins (BLTPs) are an evolutionarily conserved family of proteins that localize to membrane-contact sites and are thought to mediate the bulk transfer of lipids from a donor membrane, typically the endoplasmic reticulum, to an acceptor membrane, such as that of the cell or an organelle. Although BLTPs are fundamentally important for a wide array of cellular functions, their architecture, composition and lipid-transfer mechanisms remain poorly characterized. Here we present the subunit composition and the cryogenic electron microscopy structure of the native LPD-3 BLTP complex isolated from transgenic Caenorhabditis elegans. LPD-3 folds into an elongated, rod-shaped tunnel of which the interior is filled with ordered lipid molecules that are coordinated by a track of ionizable residues that line one side of the tunnel. LPD-3 forms a complex with two previously uncharacterized proteins, one of which we have named Spigot and the other of which remains unnamed. Spigot interacts with the N-terminal end of LPD-3 where lipids are expected to enter the tunnel, and experiments in multiple model systems indicate that Spigot has a conserved role in BLTP function. Our LPD-3 complex structural data reveal protein-lipid interactions that suggest a model for how the native LPD-3 complex mediates bulk lipid transport and provides a foundation for mechanistic studies of BLTPs. #1: ジャーナル: Nat.Struct.Mol.Biol. / 年: 2025タイトル: TMEM170 proteins are lipid scramblases associated with bridge-type lipid transporters BLTP1/Csf1 著者: Rocha-Roa C / Chandran Blair P / Sidhu G / Alvarez D / Davey M / Conibear E / Vanni S | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 添付画像 |
|---|

- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_45399.map.gz | 826.5 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-45399-v30.xmlemd-45399.xml | 31.7 KB 31.7 KB | 表示 表示 | EMDBヘッダ |
| FSC (解像度算出) | emd_45399_fsc.xml | 28.2 KB | 表示 | FSCデータファイル |
| 画像 |  emd_45399.png emd_45399.png | 77.4 KB | ||
| Filedesc metadata | emd-45399.cif.gz | 10.1 KB | ||
| その他 | emd_45399_half_map_1.map.gzemd_45399_half_map_2.map.gz | 1.5 GB 1.5 GB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-45399ftp://ftp.pdbj.org/pub/emdb/structures/EMD-45399 http://ftp.pdbj.org/pub/emdb/structures/EMD-45399ftp://ftp.pdbj.org/pub/emdb/structures/EMD-45399 | HTTPS FTP |
-関連構造データ

-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|
-マップ
| ファイル | ダウンロード / ファイル: emd_45399.map.gz / 形式: CCP4 / 大きさ: 1.6 GB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 | 画像のコントロール
画像は Spider により作成 | ||||||||||||||||||||||||||||||||||||
| ボクセルのサイズ | X=Y=Z: 0.788 Å | ||||||||||||||||||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||||||||||||||||||
| 詳細 | EMDB XML:
|
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-添付データ
-ハーフマップ: #2
| ファイル | emd_45399_half_map_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |








-ハーフマップ: #1
| ファイル | emd_45399_half_map_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |








- 試料の構成要素
試料の構成要素
-全体 : LPD-3 complex
| 全体 | 名称: LPD-3 complex |
|---|---|
| 要素 |
|
-超分子 #1: LPD-3 complex
| 超分子 | 名称: LPD-3 complex / タイプ: complex / ID: 1 / 親要素: 0 / 含まれる分子: #1-#2 |
|---|---|
| 由来(天然) | 生物種: |
-分子 #1: Bridge-like lipid transfer protein family member 1 C-terminal dom...
| 分子 | 名称: Bridge-like lipid transfer protein family member 1 C-terminal domain-containing protein タイプ: protein_or_peptide / ID: 1 / コピー数: 1 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: |
| 分子量 | 理論値: 483.608094 KDa |
| 配列 | 文字列: MSDFDIQIKI DDAQLDLKGV SFWVTASVVT LFLAWSTFVV LFFSRVSALF FTFVIDKYLR LSKNGIHFKI GGISISGLHA GKIMFRNVI YDNGDMTIKV NDGHLLFKYW KSVEHRHLNL STKRASRLHL VLNGLHVNIY NNLTKYTEIA RIRRFDWFFE N TNMNDARR ...文字列: MSDFDIQIKI DDAQLDLKGV SFWVTASVVT LFLAWSTFVV LFFSRVSALF FTFVIDKYLR LSKNGIHFKI GGISISGLHA GKIMFRNVI YDNGDMTIKV NDGHLLFKYW KSVEHRHLNL STKRASRLHL VLNGLHVNIY NNLTKYTEIA RIRRFDWFFE N TNMNDARR PQTKPPDTCR SPPSSVWENM WNLLGIVHIE VSAGCILVGN KFLPYALWTR FENLNSKTSV TESANDRALL TF EGETENV AVSLIKNEQF DFTAKDKDPP RTMGNDGCPL LQSASLEFVY KQDLLGYVTD DEPQSITLKL PLWSSEWRFG NNT VLSYGP WAEQQRFLIY SFFYPPDFQN STATAMPTRG KKRIHVKHDV KIILTKETCM DIWFMRGEQL ESIRTRCGPL SSLD MSILW ITTEKGFYWN MKAEFLNFEA TTSLIFTKLF SCKKFNVDGS FVYPLTWNGE QTWTIDYAFT KANAWFVWDH KRLFT DLIN DWIGDDPSDI SKFVPFRVHN RMKVVDGFEV IMLLNESNWV DTADMNAENV EVAIVGEKLS FECELPFVDF LPQTQM VKY EMRGEKSVAM RAKFPPDSAT APIRAALSRL ARCNSYAPPS KHGTHSLDTD VWFELWRTEL VKMDFDHHYR PLIVKSN IP SDIPFSILSD YLPPPANHPW DLEPDYLGVD ILIEGSDVKF TGLLVKLLFE LKNNYFGWYD SMTSVDDEKI DDPIKLKA S FDKTNANGMK PVEYFRTMNV DVTVRVCNVR AEMLLYSPAI DEGAEPEKVP VVFVEEVAVE VKKTKTQALI QVGVSPACA YLDKSSQGSG PGCITLSGFQ FRGHAMYSAK EVAWNMGLVE YGWIMEILVG DIAGTLDFPA HAHVLHQIME SLLMFVISPD DATKVPDRM QFCQHGQLIK ACSIAGKKTN EILGPCKTEE QMKYRQIRIS VDSVNLTFVE EKTILQISAD PVRVTICNAH E SRFTEHVC IRVPGISIRQ AVRIKEKPEN IWIEGANAAI EGVSLDIELP TPKSASPTIG KERLEFVRMH DADTKRLHFL WA DHSVWGC ACFGNTCFFG DVDEIGSTFM ETLTKKKFFV PGIERNPEKQ PQVMQSVILK NKPILSNQPH MFYKKPKNAD VVI TIRKES TGDTESFHSA RSQQSPGLRI LQSMEMSSSY ATFVDNVRVE LPSAITVPQF GEPGAILEWC QAHQATRIIN DVNT SGVNE VRFLSKPKKS QDIEYNTSRD TLGKRRLAIN GVAATSLDLF VTPIGIEAFE RLVTAASHSV PAINPCILVH MCYRD CVLK KHRQPLTESL FADEDNDSEP ISEVDITVDL PRVSIGLFQC GVKKNIVKSN HTDHITANMG LLLIDRAFIQ SKLIPA ESV SQDFSADTSN LSSTLYQLNG SAITVQLLQL TNRDAPDFGS SGTATTPNNW EHCAISRRMN NLEPRVMMDF NVSDTLI IL ERRPIILLLP DKSTTAITPI HSPANAPTPT AMNRTPTLTL TPSAGAGGGE RAEPMRKKKK MICEHYLKAD IGSVTTAL V MARPQELTAG DEFPIYEALA PVMVSWLSVV ENFLRTVDKF IHTVECWKSV AMAKVLKLAL DSTDEKVVVK VGKNRMGRT RVLSAHQASC PSCILLKTLF RWFAYAGNAP GAINHRLDIR PEFEIEETRK TALMALLSHW QSDVGKELKL VSYEDAHRFK VTRPDEAAI VALTKSKRLK RKMLEKKESS KKETRVVMEV KPEQPKAKRG RMSPAPLLKK LRKKAGDDDF DDDSMKFLSD V EMQEFNTL PLYEDYEDDE MLENLDSEPK IDDKVDLYTW MRNAQRESTL RRRKLAGGAE GSVKDDLNLK GYINPMDIQQ KA YYYNIYR WAQLQWTSLD GIEKDHWHLD YSVTLREVDV RMMAKSIKNS SDHLRQYITP AQQKVMQVRN AAVNGGMVWK MER DERRKI PLHGQWNISY SGNVEGIRFL IGMATVSLGK ELSLVLRVAM EAKNELRMHS TAESTPRNEV KVFKPVVPNQ YDLA VEWDE KVLDMTRDYE KHMQRMRTNK EDVVEKVKVM VNGSAMVSSI VLESVLNDLY VSVTISQIVL AHSKNPMPDI PVVVH AVTL STTPTAAAAE DKKTATLTKK SVSSTFKIDD LTVSLTKMKL TLSEADSSNK KSDILRCTLN SSSFNVHTNL KTLTSA KES NRPKNNLINS NIATTATLRL GALEGTMPMA AYSLHDVVMR HGKELEQQLN RLAAQPASTP LSSSTPFPSA EQSLLAK VA DMKTAPEPVV ITQAEFKPLT TLPATAAHVQ DAKGQIVRRV PVAVVSFSIE LTSIEMNIQL LPSLQAKYRI NRATSNGI T GVQANWSILL DEHFFEFCVT GQGGKTETFR LQLPSVTSDG LYQAEQGVSS QKPSTDKKLI YREGGSLQMT VVLGRVNHI FTTELLNQLM FAEHSFRTEL TALINRIRSS SFASTNSSRS AQSTNDRVNS TANLKLLLPV QTTSTPVHIE KPPLLFSIKI QTKEIPRKD EKTDKDAKTP KASGASGNLD QSQHPSHSTH VKSTPWLQLT AATPTQTAVR LTVDSLEGEL TNKWVVKEEG S KERIYGNA VIHFNAKLGQ LIKPVPTGDS VAATDVTDLQ EFATFMTQVR VENKERNMFN SSYSYHISLN RPIFLVKAAA ID KAILLWL NYKNTYDYWR NEREKVVQEK TTTKLSNAGM FSPTQIAEDA DMNLSLAINN GMYMCMPLYS HDVTEGMPAL VLS LQKSNL SVLVKKELTC KASFNGFKCS FVDDFDEQAL TQSFLDATHS DQSNCIFFPE GTYQLCSKAE ATKGPAKWVL SVSA EMQGV EIDLDTRIGK LAKLLVNTFS MIRTDDDDDM SFWGDEGELD SDEEKVEGAS ELKKLKAEEK VPWMENKMHE HSRAV FELA ARGVSNKLIE AEKHKLRQYE LIRFKAFRRN VVEKLKKGTT ASRQHTETPP PQPRPDTTSR RNSRTTSTSQ KNSEDL TTP GDIETVNFNL DVKVNITSGT CTLRTQKKEG ANQLALPGIL KRLNLGTKDI KAMFEPQIIT TTTFSIPSVE IKAYHVS DP SNRSTDEFCK DKREKISKGL AKDADKLHRD LHNKSRFGNT YINGGGGPKT STVPPPPKRG CFYIFVGLAS MPSETVVT P HLATYFEQVL EPLPPSAVFQ SQNNTREASV PDDGKGDANN EVHNIMAMDT AAFPIDFVFY LDVQSSTIRF DGKQPTSRS QTQADCLLTL PRLTLELTSK RTRDNIDNYV GGIHISGQFK GFMLKIYNPL ELEPDSSRAL QLSLDLLSFV ISRNKNSSTE PDNRVRFVF SSQISKASFE YNFRRLGELI QFPKPWYRAA IARRVFFGDQ AAPRQKDDAS DITGTTRSRL PTDPKSLQPP A ASTASTGS GSFVPHQRKP WTALVLAAIQ WNEFEVTAFM SNTMGKTTWK ATKGLVWGDA KLNSLNERDV SISFVLGSSE LC ARDGAIS GTIMLNNLKV SADHSLSADV KRVPVNKAKI RLEWITANIE WMSRRVLIAK WCGPSFKVND YYKGLKEGDH FAL SELGMN VQASWKDLQV VITKSTVDDV AAIVNRLISF IDEQLKNSRI LLGNLSASTN LKKQAQALIE SRKPTTHFWE KVLD YMSEM QMNEQLMGLM EREGAKVGGH IELKAGGISL VMMKGDMNAD TWAVFHLRDA CILFDPEARM DFLDNSSQQK IGILL KQTF CLQLGSRHGN QTENRANVCR VQTRFNNSRH LQKAEDILEF FIGDVMKIIG SADHSEKKKL KEVEVIQSPI SENENT AKS PTSTFSRFRS PGTSKTKESG PATNHNVMEL FQFPGLEAKM SSQQLNGVDD GDKYESVFQM PMDVLTTFVC DFFSEVA IE TNFNAQVSFL PELLKSYLKE SHSGTSSSHS TNSSPAVSSS KESVVSETSK DPRIFTCQEW KVEPRVRFID RIKWTPPV L DDILKKLQIF DHRNTIPKVI QRAVLDPLDA TLAASVIATL QIVDNKKTIQ KFKKSRTDSM APTPKRRDSR RSSEEVSVS IDIPDIITDI SDASFRPKHN GSGVSKGEEL FTGVVPILVE LDGDVNGHKF SVSGEGEGDA TYGKLTLKLI CTTGKLPVPW PTLVTTLGY GLMCFARYPD HMKQHDFFKS AMPEGYVQER TIFFKDDGNY KTRAEVKFEG DTLVNRIELK GIDFKEDGNI L GHKLEYNY NSHNVYITAD KQKNGIKANF KIRHNIEDGG VQLADHYQQN TPIGDGPVLL PDNHYLSYQS KLSKDPNEKR DH MVLLEFV TAAGITLGMD ELYKGSGLEV LFQGPANSGV DYKDHDGDYK DHDIDYKDDD DK UniProtKB: Bridge-like lipid transfer protein family member 1 |
-分子 #2: Defect at low temperature protein 1
| 分子 | 名称: Defect at low temperature protein 1 / タイプ: protein_or_peptide / ID: 2 / コピー数: 1 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: |
| 分子量 | 理論値: 31.56448 KDa |
| 配列 | 文字列: MRIFVFDGTS VIYVCAVIIL ILIWVAIIGQ RQIWRHRHNV ARVRPQVSLS SRISSKKAVL LRETQLDTVM RLRCENQARL TDCISLQFH GEKPYVHRMI AVDEVTLEID GQLNRIEGAV QRQAGESTYS YLKRIREKVP SIPLNLVHRI AFLQESARFR P EKFDVEQV ...文字列: MRIFVFDGTS VIYVCAVIIL ILIWVAIIGQ RQIWRHRHNV ARVRPQVSLS SRISSKKAVL LRETQLDTVM RLRCENQARL TDCISLQFH GEKPYVHRMI AVDEVTLEID GQLNRIEGAV QRQAGESTYS YLKRIREKVP SIPLNLVHRI AFLQESARFR P EKFDVEQV MELRSLLNQF LRILSAEYDS LSDEPDMAAP RGVIASFYQF GQKIMPNNSG SKRRRNKFGG SDGVRLLMKE RE EQLSLLS PLARASADSP APAAHLRRQS DSHSALLPQ UniProtKB: Lipid transport auxiliary protein 1 |
-分子 #3: Vesicle transport protein
| 分子 | 名称: Vesicle transport protein / タイプ: protein_or_peptide / ID: 3 / コピー数: 1 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: |
| 分子量 | 理論値: 16.205293 KDa |
| 配列 | 文字列: MPENKEIITI FENSRNYTNL TSESTYSLAG ALLLSGKYMN DWWEITLSIF LWMSLSFMVI SLGATILSLF TLRKHPYVCF IPIPFIIMM FIIPFVFGAP TSMVLALAMY ASKNAVSTWY CAIMGIIQTL LIFVTSVTRI HATL UniProtKB: Vesicle transport protein |
-分子 #4: 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine
| 分子 | 名称: 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine / タイプ: ligand / ID: 4 / コピー数: 18 / 式: PEE |
|---|---|
| 由来(天然) | 生物種: |
| 分子量 | 理論値: 744.034 Da |
| Chemical component information | 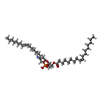 ChemComp-PEE: |
-分子 #5: HEXADECANE
| 分子 | 名称: HEXADECANE / タイプ: ligand / ID: 5 / コピー数: 12 / 式: R16 |
|---|---|
| 分子量 | 理論値: 226.441 Da |
| Chemical component information |  ChemComp-R16: |
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | particle |
-試料調製
| 緩衝液 | pH: 8 |
|---|---|
| 凍結 | 凍結剤: ETHANE |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | TFS KRIOS |
|---|---|
| 撮影 | フィルム・検出器のモデル: FEI FALCON IV (4k x 4k) 平均電子線量: 50.0 e/Å2 |
| 電子線 | 加速電圧: 300 kV / 電子線源:  FIELD EMISSION GUN FIELD EMISSION GUN |
| 電子光学系 | 照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELD / 最大 デフォーカス(公称値): 2.5 µm / 最小 デフォーカス(公称値): 1.5 µm |
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |

