ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 |  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
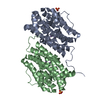
| タイトル | Class 31 model for combined refinement of Bacillus subtilis ribonucleotide reductase complex | |||||||||
 マップデータ マップデータ | ||||||||||
 試料 試料 |
| |||||||||
 キーワード キーワード | Ribonucleotide Reductase / OXIDOREDUCTASE | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報ribonucleoside-diphosphate reductase complex / ribonucleoside-diphosphate reductase / ribonucleoside-diphosphate reductase activity, thioredoxin disulfide as acceptor / deoxyribonucleotide biosynthetic process / ATP binding / metal ion binding 類似検索 - 分子機能 | |||||||||
| 生物種 |  | |||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.99 Å | |||||||||
 データ登録者 データ登録者 | Xu D / Thomas WC / Burnim AA / Ando N | |||||||||
| 資金援助 |  米国, 2件 米国, 2件
| |||||||||
 引用 引用 | ジャーナル: Nat Commun / 年: 2025 タイトル: Conformational landscapes of a class I ribonucleotide reductase complex during turnover reveal intrinsic dynamics and asymmetry. 著者: Da Xu / William C Thomas / Audrey A Burnim / Nozomi Ando / 要旨: Understanding the structural dynamics associated with enzymatic catalysis has been a long-standing goal of biochemistry. With the advent of modern cryo-electron microscopy (cryo-EM), it has become ...Understanding the structural dynamics associated with enzymatic catalysis has been a long-standing goal of biochemistry. With the advent of modern cryo-electron microscopy (cryo-EM), it has become conceivable to redefine a protein's structure as the continuum of all conformations and their distributions. However, capturing and interpreting this information remains challenging. Here, we use classification and deep-learning-based analyses to characterize the conformational heterogeneity of a class I ribonucleotide reductase (RNR) during turnover. By converting the resulting information into physically interpretable 2D conformational landscapes, we demonstrate that RNR continuously samples a wide range of motions while maintaining surprising asymmetry to regulate the two halves of its turnover cycle. Remarkably, we directly observe the appearance of highly transient conformations needed for catalysis, as well as the interaction of RNR with its endogenous reductant thioredoxin also contributing to the asymmetry and dynamics of the enzyme complex. Overall, this work highlights the role of conformational dynamics in regulating key steps in enzyme mechanisms. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
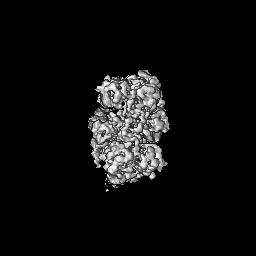
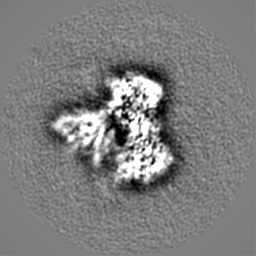
| 添付画像 |
|---|

- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_45068.map.gz | 50.1 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-45068-v30.xmlemd-45068.xml | 21.4 KB 21.4 KB | 表示 表示 | EMDBヘッダ |
| 画像 |  emd_45068.png emd_45068.png | 74.7 KB | ||
| Filedesc metadata | emd-45068.cif.gz | 7 KB | ||
| その他 | emd_45068_half_map_1.map.gzemd_45068_half_map_2.map.gz | 50 MB 50.1 MB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-45068ftp://ftp.pdbj.org/pub/emdb/structures/EMD-45068 http://ftp.pdbj.org/pub/emdb/structures/EMD-45068ftp://ftp.pdbj.org/pub/emdb/structures/EMD-45068 | HTTPS FTP |
-検証レポート
| 文書・要旨 | emd_45068_validation.pdf.gz | 1008 KB | 表示 | EMDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | emd_45068_full_validation.pdf.gz | 1007.6 KB | 表示 | |
| XML形式データ | emd_45068_validation.xml.gz | 12.1 KB | 表示 | |
| CIF形式データ | emd_45068_validation.cif.gz | 14.4 KB | 表示 | |
| アーカイブディレクトリ | https://ftp.pdbj.org/pub/emdb/validation_reports/EMD-45068ftp://ftp.pdbj.org/pub/emdb/validation_reports/EMD-45068 | HTTPS FTP |
-関連構造データ
| 関連構造データ |  9bziMC  9bw3C  9bwxC  9bx2C  9bx3C  9bx6C  9bx8C  9bx9C  9bxcC  9bxsC  9bxtC  9bxxC  9bxzC  9by0C  9by1C  9by2C  9by3C  9by7C  9by8C  9by9C  9byaC  9bycC  9bydC  9bygC  9byhC  9bylC  9bytC  9byvC  9bywC  9byxC  9byyC  9byzC  9bz2C  9bz3C  9bz5C  9bz6C  9bz9C  9bzaC  9bzdC 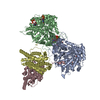 9bzeC  9bzfC  9bzhC  9bzjC 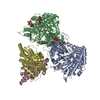 9bzkC  9bzmC  9bzoC M: このマップから作成された原子モデル C: 同じ文献を引用 ( |
|---|---|
| 類似構造データ |


























-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| 「今月の分子」の関連する項目 |
-マップ
| ファイル | ダウンロード / ファイル: emd_45068.map.gz / 形式: CCP4 / 大きさ: 64 MB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
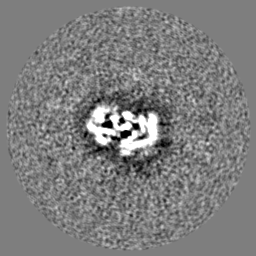
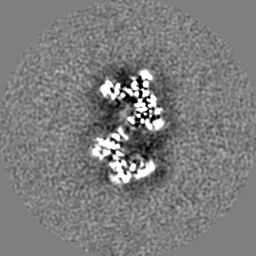
| 投影像・断面図 | 画像のコントロール
画像は Spider により作成 | ||||||||||||||||||||||||||||||||||||
| ボクセルのサイズ | X=Y=Z: 1.014 Å | ||||||||||||||||||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||||||||||||||||||
| 詳細 | EMDB XML:
|
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)
















-添付データ
-ハーフマップ: #1
| ファイル | emd_45068_half_map_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |







-ハーフマップ: #2
| ファイル | emd_45068_half_map_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |





- 試料の構成要素
試料の構成要素
-全体 : Bacillus subtilis ribonucleotide reductase complex
| 全体 | 名称: Bacillus subtilis ribonucleotide reductase complex |
|---|---|
| 要素 |
|
-超分子 #1: Bacillus subtilis ribonucleotide reductase complex
| 超分子 | 名称: Bacillus subtilis ribonucleotide reductase complex / タイプ: complex / ID: 1 / 親要素: 0 / 含まれる分子: #1-#2 |
|---|---|
| 由来(天然) | 生物種: |
-分子 #1: Ribonucleoside-diphosphate reductase subunit alpha
| 分子 | 名称: Ribonucleoside-diphosphate reductase subunit alpha / タイプ: protein_or_peptide / ID: 1 / コピー数: 2 / 光学異性体: LEVO / EC番号: ribonucleoside-diphosphate reductase |
|---|---|
| 由来(天然) | 生物種: |
| 分子量 | 理論値: 80.791469 KDa |
| 組換発現 | 生物種: |
| 配列 | 文字列: MSQNQVPKWI QLNNEIMIQK DGKFQFDKDK EAVHSYFVDY INQNTVFFHN LKEKLDYLVE NQYYEEEFLS LYSFEDIKEV FKTAYAKKF RFPSFMSAFK FYNDYALKTN DKKKILERYE DRISIVALFF ANGDTEKAKE YVNLMINQEY QPSTPTFLNA G RKRRGELV ...文字列: MSQNQVPKWI QLNNEIMIQK DGKFQFDKDK EAVHSYFVDY INQNTVFFHN LKEKLDYLVE NQYYEEEFLS LYSFEDIKEV FKTAYAKKF RFPSFMSAFK FYNDYALKTN DKKKILERYE DRISIVALFF ANGDTEKAKE YVNLMINQEY QPSTPTFLNA G RKRRGELV SCFLLEVNDS LNDISRAIDI SMQLSKLGGG VSLNLSKLRA KGEAIKDVEN ATKGVVGVMK LLDNAFRYAD QM GQRQGSG AAYLNIFHRD INDFLDTKKI SADEDVRVKT LSIGVVIPDK FVELAREDKA AYVFYPHTIY KEYGQHMDEM DMN EMYDKF VDNPRVKKEK INPRKLLEKL AMLRSESGYP YIMFQDNVNK VHANNHISKV KFSNLCSEVL QASQVSSYTD YDEE DEIGL DISCNLGSLN ILNVMEHKSI EKTVKLATDS LTHVSETTDI RNAPAVRRAN KAMKSIGLGA MNLHGYLAQN GIAYE SPEA RDFANTFFMM VNFYSIQRSA EIAKEKGETF DQYEGSTYAT GEYFDKYVST DFSPKYEKIA NLFEGMHIPT TEDWKK LKA FVAEHGMYHS YRLCIAPTGS ISYVQSSTAS VMPIMERIEE RTYGNSKTYY PMPGLASNNW FFYKEAYDMD MFKVVDM IA TIQQHIDQGI SFTLFLKDTM TTRDLNRIDL YAHHRGIKTI YYARTKDTGQ DSCLSCVV UniProtKB: Ribonucleoside-diphosphate reductase subunit alpha |
-分子 #2: Ribonucleoside-diphosphate reductase subunit beta
| 分子 | 名称: Ribonucleoside-diphosphate reductase subunit beta / タイプ: protein_or_peptide / ID: 2 / コピー数: 2 / 光学異性体: LEVO / EC番号: ribonucleoside-diphosphate reductase |
|---|---|
| 由来(天然) | 生物種: |
| 分子量 | 理論値: 40.720992 KDa |
| 組換発現 | 生物種: |
| 配列 | 文字列: MGSSHHHHHH SSGLVPRGSH MMTKIYDAAN WSKHEDDFTQ MFYNQNVKQF WLPEEIALNG DLLTWKYLGK NEQDTYMKVL AGLTLLDTE QGNTGMPIVA EHVDGHQRKA VLNFMAMMEN AVHAKSYSNI FMTLAPTETI NEVFEWVKQN KYLQKKAQMI V GLYKAIQK ...文字列: MGSSHHHHHH SSGLVPRGSH MMTKIYDAAN WSKHEDDFTQ MFYNQNVKQF WLPEEIALNG DLLTWKYLGK NEQDTYMKVL AGLTLLDTE QGNTGMPIVA EHVDGHQRKA VLNFMAMMEN AVHAKSYSNI FMTLAPTETI NEVFEWVKQN KYLQKKAQMI V GLYKAIQK DDEISLFKAM VASVYLESFL FYSGFYYPLY FYGQGKLMQS GEIINLILRD EAIHGVYVGL LAQEIYNKQT EE KKAELRE FAIDLLNQLY ENELEYTEDL YDQVGLSHDV KKFIRYNANK ALMNLGFDPY FEEEDINPIV LNGLNTKTKS HDF FSMKGN GYKKATVEPL KDDDFYFEDE KEQI UniProtKB: Ribonucleoside-diphosphate reductase subunit beta |
-分子 #3: ADENOSINE-5'-TRIPHOSPHATE
| 分子 | 名称: ADENOSINE-5'-TRIPHOSPHATE / タイプ: ligand / ID: 3 / コピー数: 2 / 式: ATP |
|---|---|
| 分子量 | 理論値: 507.181 Da |
| Chemical component information |  ChemComp-ATP: |
-分子 #4: GUANOSINE-5'-DIPHOSPHATE
| 分子 | 名称: GUANOSINE-5'-DIPHOSPHATE / タイプ: ligand / ID: 4 / コピー数: 2 / 式: GDP |
|---|---|
| 分子量 | 理論値: 443.201 Da |
| Chemical component information | 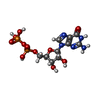 ChemComp-GDP: |
-分子 #5: THYMIDINE-5'-TRIPHOSPHATE
| 分子 | 名称: THYMIDINE-5'-TRIPHOSPHATE / タイプ: ligand / ID: 5 / コピー数: 2 / 式: TTP |
|---|---|
| 分子量 | 理論値: 482.168 Da |
| Chemical component information |  ChemComp-TTP: |
-分子 #6: MAGNESIUM ION
| 分子 | 名称: MAGNESIUM ION / タイプ: ligand / ID: 6 / コピー数: 2 / 式: MG |
|---|---|
| 分子量 | 理論値: 24.305 Da |
-分子 #7: MANGANESE (II) ION
| 分子 | 名称: MANGANESE (II) ION / タイプ: ligand / ID: 7 / コピー数: 4 / 式: MN |
|---|---|
| 分子量 | 理論値: 54.938 Da |
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | particle |
-試料調製
| 緩衝液 | pH: 7.6 詳細: 50 mM HEPES, 150 mM NaCl, 15 mM MgCl2, 5% (w/v) glycerol |
|---|---|
| グリッド | モデル: Quantifoil R1.2/1.3 / 材質: COPPER / メッシュ: 300 / 支持フィルム - 材質: CARBON / 支持フィルム - トポロジー: HOLEY / 前処理 - タイプ: GLOW DISCHARGE |
| 凍結 | 凍結剤: ETHANE / チャンバー内湿度: 100 % / チャンバー内温度: 298 K / 装置: FEI VITROBOT MARK IV |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | FEI TALOS ARCTICA |
|---|---|
| 特殊光学系 | エネルギーフィルター - スリット幅: 20 eV |
| 撮影 | フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) 検出モード: COUNTING / 平均電子線量: 50.0 e/Å2 |
| 電子線 | 加速電圧: 200 kV / 電子線源:  FIELD EMISSION GUN FIELD EMISSION GUN |
| 電子光学系 | 照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELD / 最大 デフォーカス(公称値): 2.0 µm / 最小 デフォーカス(公称値): 0.8 µm |
| 試料ステージ | 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER ホルダー冷却材: NITROGEN |
| 実験機器 |  モデル: Talos Arctica / 画像提供: FEI Company |
-画像解析
| 初期モデル | モデルのタイプ: OTHER / 詳細: Consensus map from combined refinement |
|---|---|
| 最終 再構成 | 想定した対称性 - 点群: C1 (非対称) / 解像度のタイプ: BY AUTHOR / 解像度: 3.99 Å / 解像度の算出法: FSC 0.143 CUT-OFF / ソフトウェア - 名称: RELION / 使用した粒子像数: 66000 |
| 初期 角度割当 | タイプ: MAXIMUM LIKELIHOOD / ソフトウェア - 名称: RELION |
| 最終 角度割当 | タイプ: MAXIMUM LIKELIHOOD / ソフトウェア - 名称: RELION |
-原子モデル構築 1
| 初期モデル |
| |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 精密化 | 空間: REAL / プロトコル: RIGID BODY FIT | |||||||||
| 得られたモデル | PDB-9bzi: |