- EMDB-36987: Structure of CUL3-RBX1-KLHL22 complex without CUL3 NA motif -
+
Open data
ID or keywords:
Loading...
-
Basic information
Entry
Database: EMDB / ID: EMD-36987
Title
Structure of CUL3-RBX1-KLHL22 complex without CUL3 NA motif
Map data
Sample
Complex: CUL3-RBX1-KLHL22 complex without CUL3 NA motif
Protein or peptide: Cullin-3
Protein or peptide: Kelch-like protein 22
Protein or peptide: E3 ubiquitin-protein ligase RBX1, N-terminally processed
Keywords
Cullin Ring E3 ubiquitin ligase / LIGASE
Function / homology
Function and homology information
positive regulation of mitotic cell cycle phase transition / trophectodermal cellular morphogenesis / liver morphogenesis / POZ domain binding / polar microtubule / regulation protein catabolic process at postsynapse / anaphase-promoting complex-dependent catabolic process / nuclear protein quality control by the ubiquitin-proteasome system / COPII vesicle coat assembly / cellular response to L-leucine ...positive regulation of mitotic cell cycle phase transition / trophectodermal cellular morphogenesis / liver morphogenesis / POZ domain binding / polar microtubule / regulation protein catabolic process at postsynapse / anaphase-promoting complex-dependent catabolic process / nuclear protein quality control by the ubiquitin-proteasome system / COPII vesicle coat assembly / cellular response to L-leucine / negative regulation of beige fat cell differentiation / RHOBTB3 ATPase cycle / positive regulation of T cell mediated immune response to tumor cell / cullin-RING-type E3 NEDD8 transferase / NEDD8 transferase activity / embryonic cleavage / cullin-RING ubiquitin ligase complex / Cul7-RING ubiquitin ligase complex / cellular response to chemical stress / Loss of Function of FBXW7 in Cancer and NOTCH1 Signaling / positive regulation of mitotic metaphase/anaphase transition / positive regulation of protein autoubiquitination / fibroblast apoptotic process / Notch binding / RNA polymerase II transcription initiation surveillance / protein neddylation / cell projection organization / negative regulation of Rho protein signal transduction / NEDD8 ligase activity / protein K27-linked ubiquitination / negative regulation of response to oxidative stress / RHOBTB1 GTPase cycle / VCB complex / Cul5-RING ubiquitin ligase complex / stem cell division / ubiquitin-ubiquitin ligase activity / mitotic metaphase chromosome alignment / ubiquitin-dependent protein catabolic process via the C-end degron rule pathway / SCF ubiquitin ligase complex / Cul2-RING ubiquitin ligase complex / stress fiber assembly / mitotic spindle assembly checkpoint signaling / Cul3-RING ubiquitin ligase complex / negative regulation of type I interferon production / positive regulation of cytokinesis / SCF-dependent proteasomal ubiquitin-dependent protein catabolic process / Prolactin receptor signaling / negative regulation of mitophagy / Cul4A-RING E3 ubiquitin ligase complex / Cul4-RING E3 ubiquitin ligase complex / Cul4B-RING E3 ubiquitin ligase complex / ubiquitin ligase complex scaffold activity / mitotic sister chromatid segregation / cullin family protein binding / endoplasmic reticulum to Golgi vesicle-mediated transport / protein monoubiquitination / sperm flagellum / RHOBTB2 GTPase cycle / intercellular bridge / protein autoubiquitination / ubiquitin-like ligase-substrate adaptor activity / site of DNA damage / signal transduction in response to DNA damage / Nuclear events stimulated by ALK signaling in cancer / protein K48-linked ubiquitination / negative regulation of insulin receptor signaling pathway / 14-3-3 protein binding / regulation of cellular response to insulin stimulus / positive regulation of TORC1 signaling / gastrulation / transcription-coupled nucleotide-excision repair / post-translational protein modification / intrinsic apoptotic signaling pathway / cyclin binding / T cell activation / positive regulation of protein ubiquitination / negative regulation of autophagy / Regulation of BACH1 activity / integrin-mediated signaling pathway / negative regulation of canonical NF-kappaB signal transduction / kidney development / cellular response to amino acid stimulus / negative regulation of canonical Wnt signaling pathway / G1/S transition of mitotic cell cycle / Degradation of DVL / Degradation of CRY and PER proteins / Degradation of GLI1 by the proteasome / Recognition of DNA damage by PCNA-containing replication complex / RING-type E3 ubiquitin transferase / GSK3B and BTRC:CUL1-mediated-degradation of NFE2L2 / Negative regulation of NOTCH4 signaling / Hedgehog 'on' state / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / Vif-mediated degradation of APOBEC3G / Degradation of GLI2 by the proteasome / GLI3 is processed to GLI3R by the proteasome / Ubiquitin-Mediated Degradation of Phosphorylated Cdc25A / protein destabilization / NOTCH1 Intracellular Domain Regulates Transcription / Evasion by RSV of host interferon responses Similarity search - Function
National Natural Science Foundation of China (NSFC)
2021YFA1300601
China
National Natural Science Foundation of China (NSFC)
82030081
China
National Natural Science Foundation of China (NSFC)
81874235
China
National Natural Science Foundation of China (NSFC)
31800626
China
Citation
Journal: Nat Commun / Year: 2024 Title: A conserved N-terminal motif of CUL3 contributes to assembly and E3 ligase activity of CRL3. Authors: Weize Wang / Ling Liang / Zonglin Dai / Peng Zuo / Shang Yu / Yishuo Lu / Dian Ding / Hongyi Chen / Hui Shan / Yan Jin / Youdong Mao / Yuxin Yin / Abstract: The CUL3-RING E3 ubiquitin ligases (CRL3s) play an essential role in response to extracellular nutrition and stress stimuli. The ubiquitin ligase function of CRL3s is activated through dimerization. ...The CUL3-RING E3 ubiquitin ligases (CRL3s) play an essential role in response to extracellular nutrition and stress stimuli. The ubiquitin ligase function of CRL3s is activated through dimerization. However, how and why such a dimeric assembly is required for its ligase activity remains elusive. Here, we report the cryo-EM structure of the dimeric CRL3 complex and reveal a conserved N-terminal motif in CUL3 that contributes to the dimerization assembly and the E3 ligase activity of CRL3. We show that deletion of the CUL3 N-terminal motif impairs dimeric assembly and the E3 ligase activity of both CRL3 and several other CRL3s. In addition, we found that the dynamics of dimeric assembly of CRL3 generates a variable ubiquitination zone, potentially facilitating substrate recognition and ubiquitination. These findings demonstrate that a CUL3 N-terminal motif participates in the assembly process and provide insights into the assembly and activation of CRL3s.
In the structure databanks used in Yorodumi, some data are registered as the other names, "COVID-19 virus" and "2019-nCoV". Here are the details of the virus and the list of structure data.
Jan 31, 2019. EMDB accession codes are about to change! (news from PDBe EMDB page)
EMDB accession codes are about to change! (news from PDBe EMDB page)
The allocation of 4 digits for EMDB accession codes will soon come to an end. Whilst these codes will remain in use, new EMDB accession codes will include an additional digit and will expand incrementally as the available range of codes is exhausted. The current 4-digit format prefixed with “EMD-” (i.e. EMD-XXXX) will advance to a 5-digit format (i.e. EMD-XXXXX), and so on. It is currently estimated that the 4-digit codes will be depleted around Spring 2019, at which point the 5-digit format will come into force.
The EM Navigator/Yorodumi systems omit the EMD- prefix.
Related info.:Q: What is EMD? / ID/Accession-code notation in Yorodumi/EM Navigator
Yorodumi is a browser for structure data from EMDB, PDB, SASBDB, etc.
This page is also the successor to EM Navigator detail page, and also detail information page/front-end page for Omokage search.
The word "yorodu" (or yorozu) is an old Japanese word meaning "ten thousand". "mi" (miru) is to see.
Related info.:EMDB / PDB / SASBDB / Comparison of 3 databanks / Yorodumi Search / Aug 31, 2016. New EM Navigator & Yorodumi / Yorodumi Papers / Jmol/JSmol / Function and homology information / Changes in new EM Navigator and Yorodumi
 Movie
Movie Controller
Controller


 Open data
Open data
 Basic information
Basic information
 Map data
Map data Sample
Sample Keywords
Keywords Function and homology information
Function and homology information Homo sapiens (human)
Homo sapiens (human) Authors
Authors China, 4 items
China, 4 items  Citation
Citation Structure visualization
Structure visualization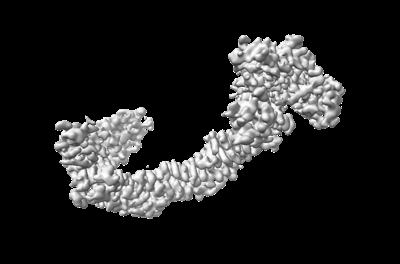
 Downloads & links
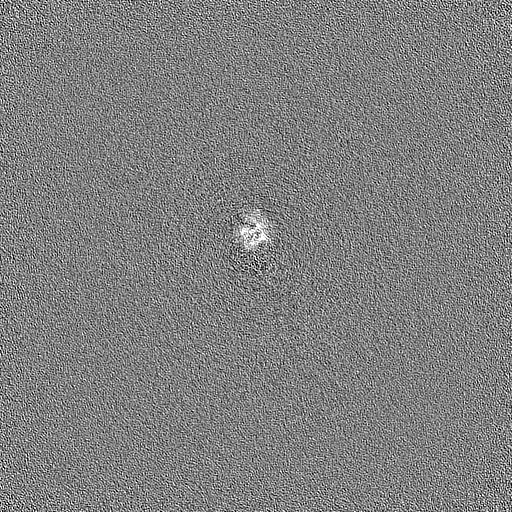
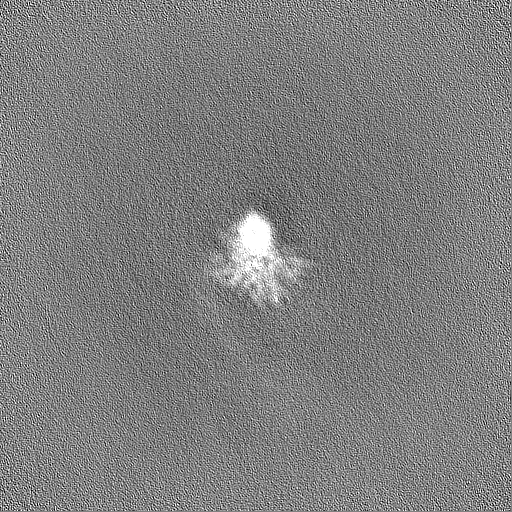
Downloads & links emd_36987.png
emd_36987.png http://ftp.pdbj.org/pub/emdb/structures/EMD-36987
http://ftp.pdbj.org/pub/emdb/structures/EMD-36987














 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





























 Sample components
Sample components
 Spodoptera frugiperda (fall armyworm)
Spodoptera frugiperda (fall armyworm) Processing
Processing Electron microscopy
Electron microscopy FIELD EMISSION GUN
FIELD EMISSION GUN

