 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry |  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Architecture of the human NALCN channelosome | |||||||||
 Map data Map data | Overall map | |||||||||
 Sample Sample |
| |||||||||
 Keywords Keywords | ion channel / channelosome / NALCN / MEMBRANE PROTEIN | |||||||||
| Function / homology |  Function and homology information Function and homology informationmonoatomic cation homeostasis / positive regulation of synaptic transmission, cholinergic / leak channel activity / viral tegument / regulation of resting membrane potential / cation channel complex / CaM pathway / Cam-PDE 1 activation / Sodium/Calcium exchangers / Calmodulin induced events ...monoatomic cation homeostasis / positive regulation of synaptic transmission, cholinergic / leak channel activity / viral tegument / regulation of resting membrane potential / cation channel complex / CaM pathway / Cam-PDE 1 activation / Sodium/Calcium exchangers / Calmodulin induced events / voltage-gated sodium channel activity / Reduction of cytosolic Ca++ levels / Activation of Ca-permeable Kainate Receptor / CREB1 phosphorylation through the activation of CaMKII/CaMKK/CaMKIV cascasde / Loss of phosphorylation of MECP2 at T308 / CREB1 phosphorylation through the activation of Adenylate Cyclase / negative regulation of high voltage-gated calcium channel activity / PKA activation / CaMK IV-mediated phosphorylation of CREB / Glycogen breakdown (glycogenolysis) / CLEC7A (Dectin-1) induces NFAT activation / Activation of RAC1 downstream of NMDARs / negative regulation of ryanodine-sensitive calcium-release channel activity / organelle localization by membrane tethering / mitochondrion-endoplasmic reticulum membrane tethering / autophagosome membrane docking / negative regulation of calcium ion export across plasma membrane / regulation of cardiac muscle cell action potential / presynaptic endocytosis / sodium channel activity / Synthesis of IP3 and IP4 in the cytosol / regulation of cell communication by electrical coupling involved in cardiac conduction / Phase 0 - rapid depolarisation / calcineurin-mediated signaling / Negative regulation of NMDA receptor-mediated neuronal transmission / Unblocking of NMDA receptors, glutamate binding and activation / RHO GTPases activate PAKs / calcium ion import across plasma membrane / Ion transport by P-type ATPases / Uptake and function of anthrax toxins / regulation of ryanodine-sensitive calcium-release channel activity / Long-term potentiation / protein phosphatase activator activity / Calcineurin activates NFAT / Regulation of MECP2 expression and activity / DARPP-32 events / monoatomic ion channel complex / Smooth Muscle Contraction / detection of calcium ion / regulation of cardiac muscle contraction / catalytic complex / RHO GTPases activate IQGAPs / regulation of cardiac muscle contraction by regulation of the release of sequestered calcium ion / monoatomic cation channel activity / calcium channel inhibitor activity / cellular response to interferon-beta / Activation of AMPK downstream of NMDARs / presynaptic cytosol / Protein methylation / regulation of release of sequestered calcium ion into cytosol by sarcoplasmic reticulum / eNOS activation / Ion homeostasis / titin binding / Tetrahydrobiopterin (BH4) synthesis, recycling, salvage and regulation / regulation of calcium-mediated signaling / sperm midpiece / voltage-gated potassium channel complex / potassium ion transmembrane transport / FCERI mediated Ca+2 mobilization / calcium channel complex / substantia nigra development / regulation of heart rate / FCGR3A-mediated IL10 synthesis / Ras activation upon Ca2+ influx through NMDA receptor / sodium ion transmembrane transport / Antigen activates B Cell Receptor (BCR) leading to generation of second messengers / calyx of Held / bioluminescence / adenylate cyclase activator activity / sarcomere / VEGFR2 mediated cell proliferation / protein serine/threonine kinase activator activity / positive regulation of synaptic transmission, GABAergic / VEGFR2 mediated vascular permeability / regulation of cytokinesis / spindle microtubule / generation of precursor metabolites and energy / positive regulation of receptor signaling pathway via JAK-STAT / Translocation of SLC2A4 (GLUT4) to the plasma membrane / calcium channel regulator activity / RAF activation / Transcriptional activation of mitochondrial biogenesis / response to calcium ion / calcium ion transmembrane transport / cellular response to type II interferon / G2/M transition of mitotic cell cycle / Stimuli-sensing channels / long-term synaptic potentiation / spindle pole / Signaling by RAF1 mutants Similarity search - Function | |||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | |||||||||
| Method | single particle reconstruction / cryo EM / Resolution: 4.5 Å | |||||||||
 Authors Authors | Wu JP / Yan Z | |||||||||
| Funding support | 1 items
| |||||||||
 Citation Citation | Journal: Cell Discov / Year: 2022 Title: Architecture of the human NALCN channelosome. Authors: Lunni Zhou / Haobin Liu / Qingqing Zhao / Jianping Wu / Zhen Yan /  Abstract: NALCN regulates the resting membrane potential by mediating the Na leak current in neurons, and it functions as a channelosome in complex with FAM155A, UNC79, and UNC80. Dysfunction of the NALCN ...NALCN regulates the resting membrane potential by mediating the Na leak current in neurons, and it functions as a channelosome in complex with FAM155A, UNC79, and UNC80. Dysfunction of the NALCN channelosome causes a broad range of neurological and developmental diseases called NALCN channelopathies in humans. How the auxiliary subunits, especially the two large components UNC79 and UNC80, assemble with NALCN and regulate its function remains unclear. Here we report an overall architecture of the human NALCN channelosome. UNC79 and UNC80 each adopt an S-shape super-helical structure consisting of HEAT and armadillo repeats, forming a super-coiled heterodimeric assembly in the cytoplasmic side, which may provide a scaffold for the binding of other potential modulators of the channelosome. The UNC79-UNC80 assembly specifically associates with the NALCN-FAM155A subcomplex through the intracellular II-III linker of NALCN. Disruptions of the interaction interfaces between UNC79 and UNC80, and between the II-III linker of NALCN and the UNC79-UNC80 assembly, significantly reduce the NALCN-mediated currents in HEK293T system, suggesting the importance of the UNC79-UNC80 assembly in regulating channelosome function. Cross-linking mass spectrometry analysis identified an additional calmodulin (CaM) bound in the carboxyl-terminal domain of NALCN. Our study thus provides a structural basis for understanding the unique assembly mechanism and functional regulation of the NALCN channelosome, and also provides an opportunity for the interpretation of many disease-related mutations in UNC80. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Supplemental images |
|---|

- Downloads & links
Downloads & links
-EMDB archive
| Map data | emd_32544.map.gz | 482.9 MB | EMDB map data format | |
|---|---|---|---|---|
| Header (meta data) | emd-32544-v30.xmlemd-32544.xml | 31.4 KB 31.4 KB | Display Display | EMDB header |
| Images |  emd_32544.png emd_32544.png | 75.3 KB | ||
| Filedesc metadata | emd-32544.cif.gz | 11.7 KB | ||
| Others | emd_32544_additional_1.map.gzemd_32544_half_map_1.map.gzemd_32544_half_map_2.map.gz | 408.3 MB 475.5 MB 475.5 MB | ||
| Archive directory |  http://ftp.pdbj.org/pub/emdb/structures/EMD-32544ftp://ftp.pdbj.org/pub/emdb/structures/EMD-32544 http://ftp.pdbj.org/pub/emdb/structures/EMD-32544ftp://ftp.pdbj.org/pub/emdb/structures/EMD-32544 | HTTPS FTP |
-Validation report
| Summary document | emd_32544_validation.pdf.gz | 821.6 KB | Display | EMDB validaton report |
|---|---|---|---|---|
| Full document | emd_32544_full_validation.pdf.gz | 821.1 KB | Display | |
| Data in XML | emd_32544_validation.xml.gz | 18.8 KB | Display | |
| Data in CIF | emd_32544_validation.cif.gz | 22.6 KB | Display | |
| Arichive directory | https://ftp.pdbj.org/pub/emdb/validation_reports/EMD-32544ftp://ftp.pdbj.org/pub/emdb/validation_reports/EMD-32544 | HTTPS FTP |
-Related structure data
| Related structure data |  7wjiMC M: atomic model generated by this map C: citing same article ( |
|---|---|
| Similar structure data |
-Links
| EMDB pages | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| Related items in Molecule of the Month |



















-Map
| File | Download / File: emd_32544.map.gz / Format: CCP4 / Size: 512 MB / Type: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Annotation | Overall map | ||||||||||||||||||||||||||||||||||||
| Projections & slices | Image control
Images are generated by Spider. | ||||||||||||||||||||||||||||||||||||
| Voxel size | X=Y=Z: 1.087 Å | ||||||||||||||||||||||||||||||||||||
| Density |
| ||||||||||||||||||||||||||||||||||||
| Symmetry | Space group: 1 | ||||||||||||||||||||||||||||||||||||
| Details | EMDB XML:
|
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-Supplemental data
-Additional map: Local map of UNC79 N-ter with UNC80 C-ter
| File | emd_32544_additional_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Annotation | Local map of UNC79 N-ter with UNC80 C-ter | ||||||||||||
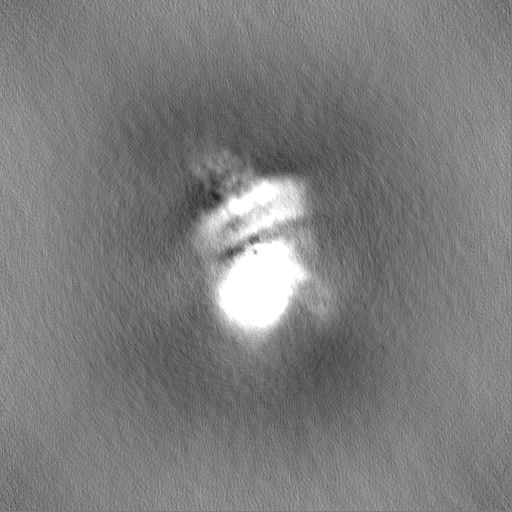
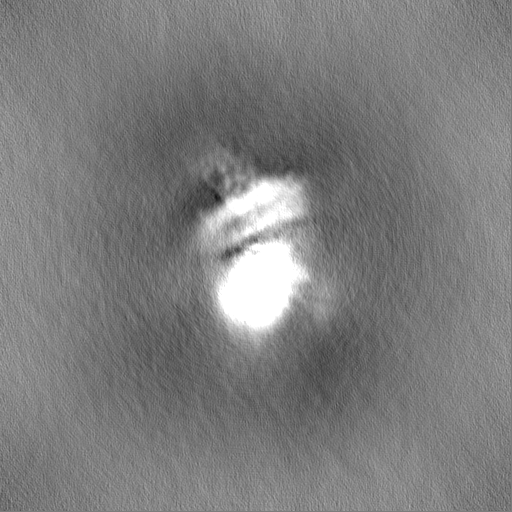
| Projections & Slices |
| ||||||||||||
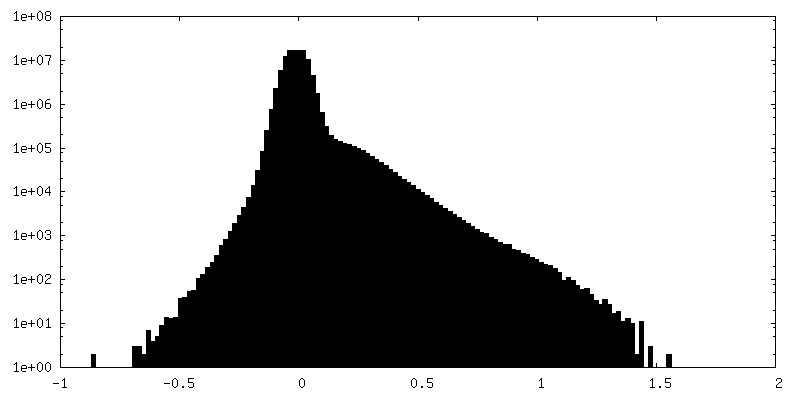
| Density Histograms |


-Half map: Half2 map of the overall map
| File | emd_32544_half_map_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Annotation | Half2 map of the overall map | ||||||||||||
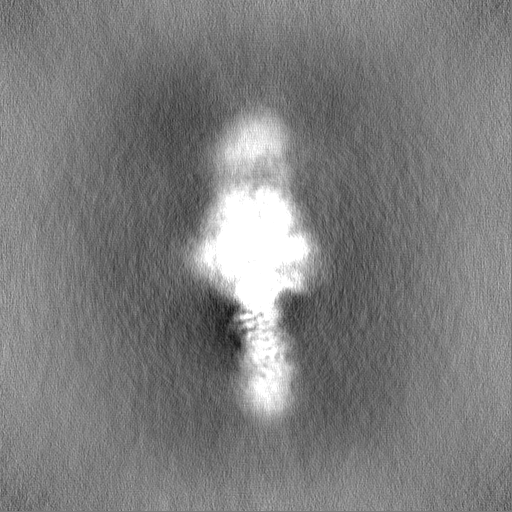
| Projections & Slices |
| ||||||||||||
| Density Histograms |

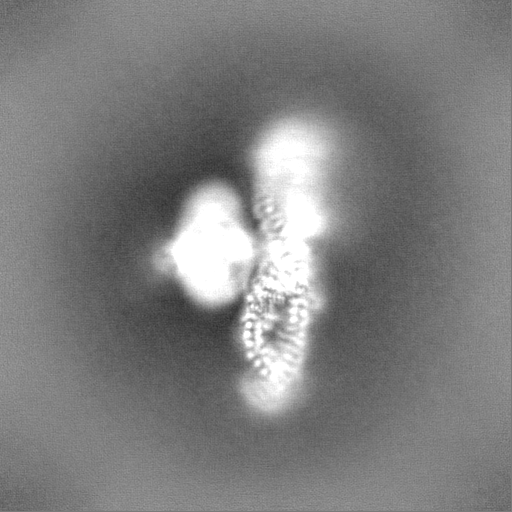
-Half map: Half1 map of the overall map
| File | emd_32544_half_map_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Annotation | Half1 map of the overall map | ||||||||||||
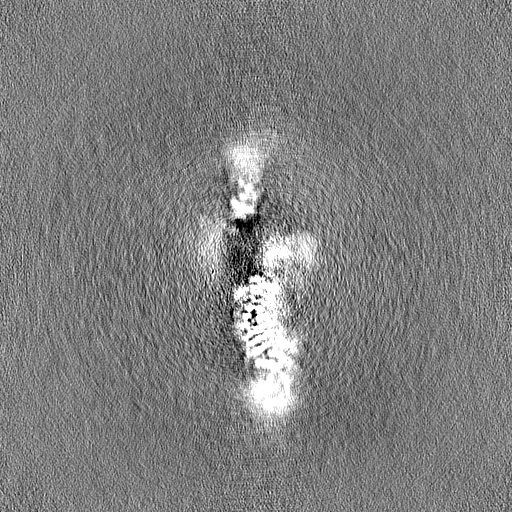
| Projections & Slices |
| ||||||||||||
| Density Histograms |




- Sample components
Sample components
-Entire : NALCN channelosome
| Entire | Name: NALCN channelosome |
|---|---|
| Components |
|
-Supramolecule #1: NALCN channelosome
| Supramolecule | Name: NALCN channelosome / type: complex / ID: 1 / Parent: 0 / Macromolecule list: all / Details: NALCN-FAM155A-UNC79-UNC80-CaM |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 900 KDa |
-Macromolecule #1: Protein unc-80 homolog
| Macromolecule | Name: Protein unc-80 homolog / type: protein_or_peptide / ID: 1 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 363.856188 KDa |
| Recombinant expression | Organism: Homo sapiens (human) |
| Sequence | String: MVKRKSSEGQ EQDGGRGIPL PIQTFLWRQT SAFLRPKLGK QYEASCVSFE RVLVENKLHG LSPALSEAIQ SISRWELVQA ALPHVLHCT ATLLSNRNKL GHQDKLGVAE TKLLHTLHWM LLEAPQDCNN ERFGGTDRGS SWGGSSSAFI HQVENQGSPG Q PCQSSSND ...String: MVKRKSSEGQ EQDGGRGIPL PIQTFLWRQT SAFLRPKLGK QYEASCVSFE RVLVENKLHG LSPALSEAIQ SISRWELVQA ALPHVLHCT ATLLSNRNKL GHQDKLGVAE TKLLHTLHWM LLEAPQDCNN ERFGGTDRGS SWGGSSSAFI HQVENQGSPG Q PCQSSSND EEENNRRKIF QNSMATVELF VFLFAPLVHR IKESDLTFRL ASGLVIWQPM WEHRQPGVSG FTALVKPIRN II TAKRSSP INSQSRTCES PNQDARHLEG LQVVCETFQS DSISPKATIS GCHRGNSFDG SLSSQTSQER GPSHSRASLV IPP CQRSRY ATYFDVAVLR CLLQPHWSEE GTQWSLMYYL QRLRHMLEEK PEKPPEPDIP LLPRPRSSSM VAAAPSLVNT HKTQ DLTMK CNEEEKSLSS EAFSKVSLTN LRRSAVPDLS SDLGMNIFKK FKSRKEDRER KGSIPFHHTG KRRPRRMGVP FLLHE DHLD VSPTRSTFSF GSFSGLGEDR RGIEKGGWQT TILGKLTRRG SSDAATEMES LSARHSHSHH TLVSDLPDPS NSHGEN TVK EVRSQISTIT VATFNTTLAS FNVGYADFFN EHMRKLCNQV PIPEMPHEPL ACANLPRSLT DSCINYSYLE DTEHIDG TN NFVHKNGMLD LSVVLKAVYL VLNHDISSRI CDVALNIVEC LLQLGVVPCV EKNRKKSENK ENETLEKRPS EGAFQFKG V SGSSTCGFGG PAVSGAGDGG GEEGGGGDGG GGGGDGGGGG GGGGGPYEKN DKNQEKDEST PVSNHRLALT MLIKIVKSL GCAYGCGEGH RGLSGDRLRH QVFRENAQNC LTKLYKLDKM QFRQTMRDYV NKDSLNNVVD FLHALLGFCM EPVTDNKAGF GNNFTTVDN KSTAQNVEGI IVSAMFKSLI TRCASTTHEL HSPENLGLYC DIRQLVQFIK EAHGNVFRRV ALSALLDSAE K LAPGKKVE ENEQESKPAG SKRSEAGSIV DKGQVSSAPE ECRSFMSGRP SQTPEHDEQM QGANLGRKDF WRKMFKSQSA AS DTSSQSE QDTSECTTAH SGTTSDRRAR SRSRRISLRK KLKLPIGKRN WLKRSSLSGL ADGVEDLLDI SSVDRLSFIR QSS KVKFTS AVKLSEGGPG SGMENGRDEE ENFFKRLGCH SFDDHLSPNQ DGGKSKNVVN LGAIRQGMKR FQFLLNCCEP GTIP DASIL AAALDLEAPV VARAALFLEC ARFVHRCNRG NWPEWMKGHH VNITKKGLSR GRSPIVGNKR NQKLQWNAAK LFYQW GDAI GVRLNELCHG ESESPANLLG LIYDEETKRR LRKEDEEEDF LDDSTVNPSK CGCPFALKMA ACQLLLEITT FLRETF SCL PRPRTEPLVD LESCRLRLDP ELDRHRYERK ISFAGVLDEN EDSKDSLHSS SHTLKSDAGV EEKKEGSPWS ASEPSIE PE GMSNAGAEEN YHRNMSWLHV MILLCNQQSF ICTHVDYCHP HCYLHHSRSC ARLVRAIKLL YGDSVDSLRE SSNISSVA L RGKKQKECSD KSCLRTPSLK KRVSDANLEG KKDSGMLKYI RLQVMSLSPA PLSLLIKAAP ILTEEMYGDI QPAAWELLL SMDEHMAGAA AAMFLLCAVK VPEAVSDMLM SEFHHPETVQ RLNAVLKFHT LWRFRYQVWP RMEEGAQQIF KIPPPSINFT LPSPVLGMP SVPMFDPPWV PQCSGSVQDP INEDQSKSFS ARAVSRSHQR AEHILKNLQQ EEEKKRLGRE ASLITAIPIT Q EACYEPTC TPNSEPEEEV EEVTNLASRR LSVSPSCTSS TSHRNYSFRR GSVWSVRSAV SAEDEEHTTE HTPNHHVPQP PQ AVFPACI CAAVLPIVHL MEDGEVREDG VAVSAVAQQV LWNCLIEDPS TVLRHFLEKL TISNRQDELM YMLRKLLLNI GDF PAQTSH ILFNYLVGLI MYFVRTPCEW GMDAISATLT FLWEVVGYVE GLFFKDLKQT MKKEQCEVKL LVTASMPGTK TLVV HGQNE CDIPTQLPVH EDTQFEALLK ECLEFFNIPE SQSTHYFLMD KRWNLIHYNK TYVRDIYPFR RSVSPQLNLV HMHPE KGQE LIQKQVFTRK LEEVGRVLFL ISLTQKIPTA HKQSHVSMLQ EDLLRLPSFP RSAIDAEFSL FSDPQAGKEL FGLDTL QKS LWIQLLEEMF LGMPSEFPWG DEIMLFLNVF NGALILHPED SALLRQYAAT VINTAVHFNH LFSLSGYQWI LPTMLQV YS DYESNPQLRQ AIEFACHQFY ILHRKPFVLQ LFASVAPLLE FPDAANNGPS KGVSAQCLFD LLQSLEGETT DILDILEL V KAEKPLKSLD FCYGNEDLTF SISEAIKLCV TVVAYAPESF RSLQMLMVLE ALVPCYLQKL KRQTSQVETV PAAREEIAA TAALATSLQA LLYSVEVLTR PMTAPQMSRC DQGHKGTTTA NHTMSSGVNT RYQEQGAKLH FIRENLHLLE EGQGIPREEL DERIAREEF RRPRESLLNI CTEFYKHCGP RLKILQNLAG EPRVIALELL DVKSHMRLAE IAHSLLKLAP YDTQTMESRG L RRYIMEML PITDWTAEAV RPALILILKR LDRMFNKIHK MPTLRRQVEW EPASNLIEGV CLTLQRQPII SFLPHLRSLI NV CVNLVMG VVGPSSVADG LPLLHLSPYL SPPLPFSTAV VRLVALQIQA LKEDFPLSHV ISPFTNQERR EGMLLNLLIP FVL TVGSGS KDSPWLEQPE VQLLLQTVIN VLLPPRIIST SRSKNFMLES SPAHCSTPGD AGKDLRREGL AESTSQAAYL ALKV ILVCF ERQLGSQWYW LSLQVKEMAL RKVGGLALWD FLDFIVRTRI PIFVLLRPFI QCKLLAQPAE NHEELSARQH IADQL ERRF IPRPLCKSSL IAEFNSELKI LKEAVHSGSA YQGKTSISTV GTSTSAYRLS LATMSRSNTG TGTVWEQDSE PSQQAS QDT LSRTDEEDEE NDSISMPSVV SEQEAYLLSA IGRRRFSSHV SSMSVPQAEV GMLPSQSEPN VLDDSQGLAA EGSLSRV AS IQSEPGQQNL LVQQPLGRKR GLRQLRRPLL SRQKTQTEPR NRQGARLSTT RRSIQPKTKP SADQKRSVTF IEAQPEPA A APTDALPATG QLQGCSPAPS RKPEAMDEPV LTSSPAIVVA DLHSVSPKQS ENFPTEEGEK EEDTEAQGAT AHSPLSAQL SDPDDFTGLE TSSLLQHGDT VLHISEENGM ENPLLSSQFT FTPTELGKTD AVLDESHV UniProtKB: Protein unc-80 homolog |
-Macromolecule #2: Protein unc-79 homolog
| Macromolecule | Name: Protein unc-79 homolog / type: protein_or_peptide / ID: 2 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 298.239656 KDa |
| Recombinant expression | Organism: Homo sapiens (human) |
| Sequence | String: MSTKAEQFAS KIRYLQEYHN RVLHNIYPVP SGTDIANTLK YFSQTLLSIL SRTGKKENQD ASNLTVPMTM CLFPVPFPLT PSLRPQVSS INPTVTRSLL YSVLRDAPSE RGPQSRDAQL SDYPSLDYQG LYVTLVTLLD LVPLLQHGQH DLGQSIFYTT T CLLPFLND ...String: MSTKAEQFAS KIRYLQEYHN RVLHNIYPVP SGTDIANTLK YFSQTLLSIL SRTGKKENQD ASNLTVPMTM CLFPVPFPLT PSLRPQVSS INPTVTRSLL YSVLRDAPSE RGPQSRDAQL SDYPSLDYQG LYVTLVTLLD LVPLLQHGQH DLGQSIFYTT T CLLPFLND DILSTLPYTM ISTLATFPPF LHKDIIEYLS TSFLPMAILG SSRREGVPAH VNLSASSMLM IAMQYTSNPV YH CQLLECL MKYKQEVWKD LLYVIAYGPS QVKPPAVQML FHYWPNLKPP GAISEYRGLQ YTAWNPIHCQ HIECHNAINK PAV KMCIDP SLSVALGDKP PPLYLCEECS ERIAGDHSEW LIDVLLPQAE ISAICQKKNC SSHVRRAVVT CFSAGCCGRH GNRP VRYCK RCHSNHHSNE VGAAAETHLY QTSPPPINTR ECGAEELVCA VEAVISLLKE AEFHAEQREH ELNRRRQLGL SSSHH SLDN ADFDNKDDDK HDQRLLSQFG IWFLVSLCTP SENTPTESLA RLVAMVFQWF HSTAYMMDDE VGSLVEKLKP QFVTKW LKT VCDVRFDVMV MCLLPKPMEF ARVGGYWDKS CSTVTQLKEG LNRILCLIPY NVINQSVWEC IMPEWLEAIR TEVPDNQ LK EFREVLSKMF DIELCPLPFS MEEMFGFISC RFTGYPSSVQ EQALLWLHVL SELDIMVPLQ LLISMFSDGV NSVKELAN Q RKSRVSELAG NLASRRVSVA SDPGRRVQHN MLSPFHSPFQ SPFRSPLRSP FRSPFKNFGH PGGRTIDFDC EDDEMNLNC FILMFDLLLK QMELQDDGIT MGLEHSLSKD IISIINNVFQ APWGGSHTCQ KDEKAIECNL CQSSILCYQL ACELLERLAP KEESRLVEP TDSLEDSLLS SRPEFIIGPE GEEEENPASK HGENPGNCTE PVEHAAVKND TERKFCYQQL PVTLRLIYTI F QEMAKFEE PDILFNMLNC LKILCLHGEC LYIARKDHPQ FLAYIQDHML IASLWRVVKS EFSQLSSLAV PLLLHALSLP HG ADIFWTI INGNFNSKDW KMRFEAVEKV AVICRFLDIH SVTKNHLLKY SLAHAFCCFL TAVEDVNPAV ATRAGLLLDT IKR PALQGL CLCLDFQFDT VVKDRPTILS KLLLLHFLKQ DIPALSWEFF VNRFETLSLE AQLHLDCNKE FPFPTTITAV RTNV ANLSD AALWKIKRAR FARNRQKSVR SLRDSVKGPV ESKRALSLPE TLTSKIRQQS PENDNTIKDL LPEDAGIDHQ TVHQL ITVL MKFMAKDESS AESDISSAKA FNTVKRHLYV LLGYDQQEGC FMIAPQKMRL STCFNAFIAG IAQVMDYNIN LGKHLL PLV VQVLKYCSCP QLRHYFQQPP RCSLWSLKPH IRQMWLKALL VILYKYPYRD CDISKILLHL IHITVNTLNA QYHSCKP HA TAGPLYSDNS NISRYSEKEK GEIELAEYRE TGALQDSLLH CVREESIPKK KLRSFKQKSL DIGNADSLLF TLDEHRRK S CIDRCDIEKP PTQAAYIAQR PNDPGRSRQN SATRPDNSEI PENPAMEGFP DARRPVIPEV RLNCMETFEV KVDSPVKPA PKEDLDLIDL SSDSTSGPEK HSILSTSDSD SLVFEPLPPL RIVESDEEEE TMNQGDDGPS GKNAASSPSV PSHPSVLSLS TAPLVQVSV EDCSKDFSSK DSGNNQSAGN TDSALITLED PMDAEGSSKP EELPEFSCGS PLTLKQKRDL LQKSFALPEM S LDDHPDPG TEGEKPGELM PSSGAKTVLL KVPEDAENPT ESEKPDTSAE SDTEQNPERK VEEDGAEESE FKIQIVPRQR KQ RKIAVSA IQREYLDISF NILDKLGEQK DPDPSTKGLS TLEMPRESSS APTLDAGVPE TSSHSSISTQ YRQMKRGSLG VLT MSQLMK RQLEHQSSAP HNISNWDTEQ IQPGKRQCNV PTCLNPDLEG QPLRMRGATK SSLLSAPSIV SMFVPAPEEF TDEQ PTVMT DKCHDCGAIL EEYDEETLGL AIVVLSTFIH LSPDLAAPLL LDIMQSVGRL ASSTTFSNQA ESMMVPGNAA GVAKQ FLRC IFHQLAPNGI FPQLFQSTIK DGTFLRTLAS SLMDFNELSS IAALSQLLEG LNNKKNLPAG GAMIRCLENI ATFMEA LPM DSPSSLWTTI SNQFQTFFAK LPCVLPLKCS LDSSLRIMIC LLKIPSTNAT RSLLEPFSKL LSFVIQNAVF TLAYLVE LC GLCYRAFTKE RDKFYLSRSV VLELLQALKL KSPLPDTNLL LLVQFICADA GTKLAESTIL SKQMIASVPG CGTAAMEC V RQYINEVLDF MADMHTLTKL KSHMKTCSQP LHEDTFGGHL KVGLAQIAAM DISRGNHRDN KAVIRYLPWL YHPPSAMQQ GPKEFIECVS HIRLLSWLLL GSLTHNAVCP NASSPCLPIP LDAGSHVADH LIVILIGFPE QSKTSVLHMC SLFHAFIFAQ LWTVYCEQS AVATNLQNQN EFSFTAILTA LEFWSRVTPS ILQLMAHNKV MVEMVCLHVI SLMEALQECN STIFVKLIPM W LPMIQSNI KHLSAGLQLR LQAIQNHVNH HSLRTLPGSG QSSAGLAALR KWLQCTQFKM AQVEIQSSEA ASQFYPLDEV DA GSDYKDD DKGSDYKDDD K UniProtKB: Protein unc-79 homolog |
-Macromolecule #3: Calmodulin-1
| Macromolecule | Name: Calmodulin-1 / type: protein_or_peptide / ID: 3 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 16.852545 KDa |
| Recombinant expression | Organism: Homo sapiens (human) |
| Sequence | String: MADQLTEEQI AEFKEAFSLF DKDGDGTITT KELGTVMRSL GQNPTEAELQ DMINEVDADG NGTIDFPEFL TMMARKMKDT DSEEEIREA FRVFDKDGNG YISAAELRHV MTNLGEKLTD EEVDEMIREA DIDGDGQVNY EEFVQMMTAK UniProtKB: Calmodulin-1 |
-Macromolecule #4: Sodium leak channel non-selective protein,Extended tegument prote...
| Macromolecule | Name: Sodium leak channel non-selective protein,Extended tegument protein pp150 type: protein_or_peptide / ID: 4 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 229.017703 KDa |
| Recombinant expression | Organism: Homo sapiens (human) |
| Sequence | String: MLKRKQSSRV EAQPVTDFGP DESLSDNADI LWINKPWVHS LLRICAIISV ISVCMNTPMT FEHYPPLQYV TFTLDTLLMF LYTAEMIAK MHIRGIVKGD SSYVKDRWCV FDGFMVFCLW VSLVLQVFEI ADIVDQMSPW GMLRIPRPLI MIRAFRIYFR F ELPRTRIT ...String: MLKRKQSSRV EAQPVTDFGP DESLSDNADI LWINKPWVHS LLRICAIISV ISVCMNTPMT FEHYPPLQYV TFTLDTLLMF LYTAEMIAK MHIRGIVKGD SSYVKDRWCV FDGFMVFCLW VSLVLQVFEI ADIVDQMSPW GMLRIPRPLI MIRAFRIYFR F ELPRTRIT NILKRSGEQI WSVSIFLLFF LLLYGILGVQ MFGTFTYHCV VNDTKPGNVT WNSLAIPDTH CSPELEEGYQ CP PGFKCMD LEDLGLSRQE LGYSGFNEIG TSIFTVYEAA SQEGWVFLMY RAIDSFPRWR SYFYFITLIF FLAWLVKNVF IAV IIETFA EIRVQFQQMW GSRSSTTSTA TTQMFHEDAA GGWQLVAVDV NKPQGRAPAC LQKMMRSSVF HMFILSMVTV DVIV AASNY YKGENFRRQY DEFYLAEVAF TVLFDLEALL KIWCLGFTGY ISSSLHKFEL LLVIGTTLHV YPDLYHSQFT YFQVL RVVR LIKISPALED FVYKIFGPGK KLGSLVVFTA SLLIVMSAIS LQMFCFVEEL DRFTTFPRAF MSMFQILTQE GWVDVM DQT LNAVGHMWAP VVAIYFILYH LFATLILLSL FVAVILDNLE LDEDLKKLKQ LKQSEANADT KEKLPLRLRI FEKFPNR PQ MVKISKLPSD FTVPKIRESF MKQFIDRQQQ DTCCLLRSLP TTSSSSCDHS KRSAIEDNKY IDQKLRKSVF SIRARNLL E KETAVTKILR ACTRQRMLSG SFEGQPAKER SILSVQHHIR QERRSLRHGS NSQRISRGKS LETLTQDHSN TVRYRNAQR EDSEIKMIQE KKEQAEMKRK VQEEELRENH PYFDKPLFIV GREHRFRNFC RVVVRARFNA SKTDPVTGAV KNTKYHQLYD LLGLVTYLD WVMIIVTICS CISMMFESPF RRVMHAPTLQ IAEYVFVIFM SIELNLKIMA DGLFFTPTAV IRDFGGVMDI F IYLVSLIF LCWMPQNVPA ESGAQLLMVL RCLRPLRIFK LVPQMRKVVR ELFSGFKEIF LVSILLLTLM LVFASFGVQL FA GKLAKCN DPNIIRREDC NGIFRINVSV SKNLNLKLRP GEKKPGFWVP RVWANPRNFN FDNVGNAMLA LFEVLSLKGW VEV RDVIIH RVGPIHGIYI HVFVFLGCMI GLTLFVGVVI ANFNENKGTA LLTVDQRRWE DLKSRLKIAQ PLHLPPRPDN DGFR AKMYD ITQHPFFKRT IALLVLAQSV LLSVKWDVED PVTVPLATMS VVFTFIFVLE VTMKIIAMSP AGFWQSRRNR YDLLV TSLG VVWVVLHFAL LNAYTYMMGA CVIVFRFFSI CGKHVTLKML LLTVVVSMYK SFFIIVGMFL LLLCYAFAGV VLFGTV KYG ENINRHANFS SAGKAITVLF RIVTGEDWNK IMHDCMVQPP FCTPDEFTYW ATDCGNYAGA LMYFCSFYVI IAYIMLN LL VAIIVENFSL FYSTEEDQLL SYNDLRHFQI IWNMVDDKRE GVIPTFRVKF LLRLLRGRLE VDLDKDKLLF KHMCYEME R LHNGGDVTFH DVLSMLSYRS VDIRKSLQLE ELLAREQLEY TIEEEVAKQT IRMWLKKCLK RIRAKQQQSC SIIHSLRES QQQELSRFLN PPSIETTQPS EDTNANSQDN SMQPETSSQQ QLLSPTLSDR GGSRQDAADA GKPQRKFGQW RLPSAPKPIS HSVSSVNLR FGGRTTMKSV VCKMNPMTDA ASCGSEVKKW WTRQLTVESD ESGDDLLDIL EGSENLYFQG GGGSMVSKGE E LFTGVVPI LVELDGDVNG HKFSVSGEGE GDATYGKLTL KFICTTGKLP VPWPTLVTTL TYGVQCFSRY PDHMKQHDFF KS AMPEGYV QERTIFFKDD GNYKTRAEVK FEGDTLVNRI ELKGIDFKED GNILGHKLEY NYNSHNVYIM ADKQKNGIKV NFK IRHNIE DGSVQLADHY QQNTPIGDGP VLLPDNHYLS TQSALSKDPN EKRDHMVLLE FVTAAGITLG MDELYK UniProtKB: Sodium leak channel NALCN, Extended tegument protein pp150 |
-Macromolecule #5: Transmembrane protein FAM155A
| Macromolecule | Name: Transmembrane protein FAM155A / type: protein_or_peptide / ID: 5 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 51.550484 KDa |
| Recombinant expression | Organism: Homo sapiens (human) |
| Sequence | String: MTRGAWMCRQ YDDGLKIWLA APRENEKPFI DSERAQKWRL SLASLLFFTV LLSDHLWFCA EAKLTRARDK EHQQQQRQQQ QQQQQQRQR QQQQQQRRQQ EPSWPALLAS MGESSPAAQA HRLLSASSSP TLPPSPGDGG GGGGKGNRGK DDRGKALFLG N SAKPVWRL ...String: MTRGAWMCRQ YDDGLKIWLA APRENEKPFI DSERAQKWRL SLASLLFFTV LLSDHLWFCA EAKLTRARDK EHQQQQRQQQ QQQQQQRQR QQQQQQRRQQ EPSWPALLAS MGESSPAAQA HRLLSASSSP TLPPSPGDGG GGGGKGNRGK DDRGKALFLG N SAKPVWRL ETCYPQGASS GQCFTVENAD AVCARNWSRG AAGGDGQEVR SKHPTPLWNL SDFYLSFCNS YTLWELFSGL SS PNTLNCS LDVVLKEGGE MTTCRQCVEA YQDYDHHAQE KYEEFESVLH KYLQSEEYSV KSCPEDCKIV YKAWLCSQYF EVT QFNCRK TIPCKQYCLE VQTRCPFILP DNDEVIYGGL SSFICTGLYE TFLTNDEPEC CDVRREEKSN NPSKGTVEKS GSCH RTSLT VSSATRLCNS RLKLCVLVLI LLHTVLTASA AQNTAGLSFG GINTLEENST NEE UniProtKB: NALCN channel auxiliary factor 1 |
-Experimental details
-Structure determination
| Method | cryo EM |
|---|---|
 Processing Processing | single particle reconstruction |
| Aggregation state | particle |
-Sample preparation
| Concentration | 0.1 mg/mL |
|---|---|
| Buffer | pH: 7.4 |
| Grid | Model: Quantifoil R1.2/1.3 / Material: COPPER / Mesh: 300 / Support film - Material: CARBON / Support film - topology: CONTINUOUS / Support film - Film thickness: 2 / Pretreatment - Type: GLOW DISCHARGE |
| Vitrification | Cryogen name: ETHANE / Chamber humidity: 100 % / Chamber temperature: 281 K |
- Electron microscopy
Electron microscopy
| Microscope | FEI TITAN KRIOS |
|---|---|
| Image recording | Film or detector model: GATAN K3 (6k x 4k) / Average electron dose: 50.0 e/Å2 |
| Electron beam | Acceleration voltage: 300 kV / Electron source:  FIELD EMISSION GUN FIELD EMISSION GUN |
| Electron optics | Illumination mode: FLOOD BEAM / Imaging mode: BRIGHT FIELD / Cs: 2.7 mm / Nominal defocus max: 2.0 µm / Nominal defocus min: 1.4000000000000001 µm / Nominal magnification: 81000 |
| Sample stage | Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER / Cooling holder cryogen: NITROGEN |
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
