 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 |  | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Structure of recombinant RyR2 mutant K4593A (Ca2+ dataset) | |||||||||||||||||||||
 マップデータ マップデータ | Structure of recombinant RyR2 mutant K4593A (Ca2 dataset) | |||||||||||||||||||||
 試料 試料 |
| |||||||||||||||||||||
 キーワード キーワード | CALCIUM / CALCIUM CHANNEL / CALCIUM TRANSPORT / ION TRANSPORT / IONIC CHANNEL / METAL TRANSPORT / ER/SR MEMBRANE / RYANODINE RECEPTOR / RYANODINE / RECEPTOR / WILD TYPE / MEMBRANE PROTEIN | |||||||||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報establishment of protein localization to endoplasmic reticulum / type B pancreatic cell apoptotic process / Purkinje myocyte to ventricular cardiac muscle cell signaling / regulation of atrial cardiac muscle cell action potential / left ventricular cardiac muscle tissue morphogenesis / suramin binding / regulation of AV node cell action potential / regulation of SA node cell action potential / Stimuli-sensing channels / regulation of ventricular cardiac muscle cell action potential ...establishment of protein localization to endoplasmic reticulum / type B pancreatic cell apoptotic process / Purkinje myocyte to ventricular cardiac muscle cell signaling / regulation of atrial cardiac muscle cell action potential / left ventricular cardiac muscle tissue morphogenesis / suramin binding / regulation of AV node cell action potential / regulation of SA node cell action potential / Stimuli-sensing channels / regulation of ventricular cardiac muscle cell action potential / ventricular cardiac muscle cell action potential / positive regulation of sequestering of calcium ion / negative regulation of calcium-mediated signaling / embryonic heart tube morphogenesis / cardiac muscle hypertrophy / Ion homeostasis / negative regulation of insulin secretion involved in cellular response to glucose stimulus / neuronal action potential propagation / negative regulation of release of sequestered calcium ion into cytosol / calcium ion transport into cytosol / insulin secretion involved in cellular response to glucose stimulus / ryanodine-sensitive calcium-release channel activity / response to caffeine / response to redox state / release of sequestered calcium ion into cytosol by sarcoplasmic reticulum / 'de novo' protein folding / negative regulation of heart rate / cellular response to caffeine / calcium ion transmembrane import into cytosol / FK506 binding / response to muscle activity / protein kinase A regulatory subunit binding / protein kinase A catalytic subunit binding / positive regulation of the force of heart contraction / intracellularly gated calcium channel activity / smooth endoplasmic reticulum / smooth muscle contraction / detection of calcium ion / T cell proliferation / regulation of cardiac muscle contraction by regulation of the release of sequestered calcium ion / positive regulation of heart rate / calcium channel inhibitor activity / Ion homeostasis / regulation of release of sequestered calcium ion into cytosol by sarcoplasmic reticulum / response to muscle stretch / release of sequestered calcium ion into cytosol / cellular response to epinephrine stimulus / calcium channel complex / sarcoplasmic reticulum membrane / regulation of heart rate / protein maturation / sarcoplasmic reticulum / sarcomere / calcium channel regulator activity / peptidylprolyl isomerase / peptidyl-prolyl cis-trans isomerase activity / establishment of localization in cell / calcium-mediated signaling / calcium ion transmembrane transport / calcium channel activity / Stimuli-sensing channels / Z disc / intracellular calcium ion homeostasis / calcium ion transport / positive regulation of cytosolic calcium ion concentration / protein refolding / transmembrane transporter binding / calmodulin binding / response to hypoxia / signaling receptor binding / calcium ion binding / protein kinase binding / enzyme binding / protein-containing complex / identical protein binding / membrane / cytoplasm 類似検索 - 分子機能 | |||||||||||||||||||||
| 生物種 |   Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||||||||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.8 Å | |||||||||||||||||||||
 データ登録者 データ登録者 | Kobayashi T / Tsutsumi A / Kurebayashi N / Kodama M / Kikkawa M / Murayama T / Ogawa H | |||||||||||||||||||||
| 資金援助 |  日本, 6件 日本, 6件
| |||||||||||||||||||||
 引用 引用 | ジャーナル: Nat Commun / 年: 2022 タイトル: Molecular basis for gating of cardiac ryanodine receptor explains the mechanisms for gain- and loss-of function mutations 著者: Kobayashi T / Tsutsumi A / Kurebayashi N / Saito K / Kodama M / Sakurai T / Kikkawa M / Murayama T / Ogawa H | |||||||||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 添付画像 |
|---|

- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_32037.map.gz | 116.6 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-32037-v30.xmlemd-32037.xml | 24.3 KB 24.3 KB | 表示 表示 | EMDBヘッダ |
| 画像 | 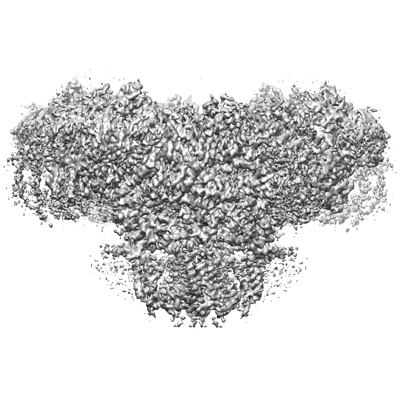 emd_32037.png emd_32037.png | 128.3 KB | ||
| Filedesc metadata | emd-32037.cif.gz | 8.8 KB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-32037ftp://ftp.pdbj.org/pub/emdb/structures/EMD-32037 http://ftp.pdbj.org/pub/emdb/structures/EMD-32037ftp://ftp.pdbj.org/pub/emdb/structures/EMD-32037 | HTTPS FTP |
-検証レポート
| 文書・要旨 | emd_32037_validation.pdf.gz | 725.5 KB | 表示 | EMDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | emd_32037_full_validation.pdf.gz | 725.1 KB | 表示 | |
| XML形式データ | emd_32037_validation.xml.gz | 6.5 KB | 表示 | |
| CIF形式データ | emd_32037_validation.cif.gz | 7.6 KB | 表示 | |
| アーカイブディレクトリ | https://ftp.pdbj.org/pub/emdb/validation_reports/EMD-32037ftp://ftp.pdbj.org/pub/emdb/validation_reports/EMD-32037 | HTTPS FTP |
-関連構造データ


-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| 「今月の分子」の関連する項目 |







-マップ
| ファイル | ダウンロード / ファイル: emd_32037.map.gz / 形式: CCP4 / 大きさ: 125 MB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | Structure of recombinant RyR2 mutant K4593A (Ca2 dataset) | ||||||||||||||||||||||||||||||||||||
| 投影像・断面図 | 画像のコントロール
画像は Spider により作成 | ||||||||||||||||||||||||||||||||||||
| ボクセルのサイズ | X=Y=Z: 1.328 Å | ||||||||||||||||||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||||||||||||||||||
| 詳細 | EMDB XML:
|
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)



















-添付データ
- 試料の構成要素
試料の構成要素
-全体 : Recombinant RyR2 mutant K4593A in the presence of Ca2+
| 全体 | 名称: Recombinant RyR2 mutant K4593A in the presence of Ca2+ |
|---|---|
| 要素 |
|
-超分子 #1: Recombinant RyR2 mutant K4593A in the presence of Ca2+
| 超分子 | 名称: Recombinant RyR2 mutant K4593A in the presence of Ca2+ タイプ: cell / ID: 1 / 親要素: 0 / 含まれる分子: #1-#2 / 詳細: in complex with FKBP12.6 |
|---|---|
| 由来(天然) | 生物種: |
-分子 #1: Ryanodine receptor 2
| 分子 | 名称: Ryanodine receptor 2 / タイプ: protein_or_peptide / ID: 1 / コピー数: 4 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: |
| 分子量 | 理論値: 533.595438 KDa |
| 組換発現 | 生物種: Homo sapiens (ヒト) |
| 配列 | 文字列: MADAGEGEDE IQFLRTDDEV VLQCTATIHK EQQKLCLAAE GFGNRLCFLE STSNSKNVPP DLSICTFVLE QSLSVRALQE MLANTVEKS EGQVDVEKWK FMMKTAQGGG HRTLLYGHAI LLRHSYSGMY LCCLSTSRSS TDKLAFDVGL QEDTTGEACW W TIHPASKQ ...文字列: MADAGEGEDE IQFLRTDDEV VLQCTATIHK EQQKLCLAAE GFGNRLCFLE STSNSKNVPP DLSICTFVLE QSLSVRALQE MLANTVEKS EGQVDVEKWK FMMKTAQGGG HRTLLYGHAI LLRHSYSGMY LCCLSTSRSS TDKLAFDVGL QEDTTGEACW W TIHPASKQ RSEGEKVRVG DDLILVSVSS ERYLHLSYGN SSWHVDAAFQ QTLWSVAPIS SGSEAAQGYL IGGDVLRLLH GH MDECLTV PSGEHGEEQR RTVHYEGGAV SVHARSLWRL ETLRVAWSGS HIRWGQPFRL RHVTTGKYLS LMEDKNLLLM DKE KADVKS TAFAFRSSKE KLDVGVRKEV DGMGTSEIKY GDSICYIQHV DTGLWLTYQA VDVKSARMGS IQRKAIMHHE GHMD DGLNL SRSQHEESRT ARVIRSTVFL FNRFIRGLDA LSKKVKLPTI DLPIESVSLS LQDLIGYFHP PDEHLEHEDK QNRLR ALKN RQNLFQEEGM INLVLECIDR LHVYSSAAHF ADVAGREAGE SWKSILNSLY ELLAALIRGN RKNCAQFSGS LDWLIS RLE RLEASSGILE VLHCVLVESP EALNIIKEGH IKSIISLLDK HGRNHKVLDV LCSLCVCHGV AVRSNQHLIC DNLLPGR DL LLQTRLVNHV SSMRPNIFLG VSEGSAQYKK WYYELMVDHT EPFVTAEATH LRVGWASTEG YSPYPGGGEE WGGNGVGD D LFSYGFDGLH LWSGCIARTV SSPNQHLLRT DDVISCCLDL SAPSISFRIN GQPVQGMFEN FNIDGLFFPV VSFSAGIKV RFLLGGRHGE FKFLPPPGYA ACYEAVLPKE KLKVEHSREY KQERTYTRDL LGPTVSLTQA AFTPVPVDTS QIVLPPHLER IRERLAENI HELWVMNKIE LGWQYGPVRD DNKRQHPCLV EFCKLPEQER NYNLQMSLET LKTLLALGCH VGIADEHAEE K VKKMKLPK NYQLTSGYKP APMDLSFIKL TPSQEAMVDK LAENAHNVWA RDRIRQGWTY GIQQDVKNRR NPRLVPYTLL DD RTKKSNK DSLREAVRTL LGYGYHLEAP DQDHASRAEV CSGTGERFRI FRAEKTYAVK AGRWYFEFEA VTAGDMRVGW SRP GCQPDL ELGSDDRAFA FDGFKAQRWH QGNEHYGRSW QAGDVVGCMV DMNEHTMMFT LNGEILLDDS GSELAFKDFD VGDG FIPVC SLGVAQVGRM NFGKDVSTLK YFTICGLQEG YEPFAVNTNR DITMWLSKRL PQFLQVPSNH EHIEVTRIDG TIDSS PCLK VTQKSFGSQN NNTDIMFYRL SMPIECA(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)DSD FEVLMKTAHG HLVPDRIDKD KETPKPEFNN HKDYAQEKPS RLKQ(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) PLSAG LFKSEHKNPV PQCPPRLHVQ FLSHVLWSRM PNQFLKVDVS RISERQGWLV QCLDPLQFMS LHIPEENRSV DILEL TEQE ELLQFHYHTL RLYSAVCALG NHRVAHALCS HVDEPQLLYA IENKYMPGLL RAGYYDLLID IHLSSYATAR LMMNNE FIV PMTEETKSIT LFPDENKKHG LPGIGLSTSL RPRMRFSSPS FVSISNDCYQ YSPEFPLDIL KAKTIQMLTE AVKEGSL HA RDPVGGTTEF LFVPLIKLFY TLLIMGIFHN EDLKHILQLI EPSVFKEAAV PEEEGGTPEK EISIEDAKLE GEEEAKGG K RPKEGLLQMK LPEPVKLQMC LLLQYLCDCQ VRHRIEAIVA FSDDFVAKLQ DNQRFRYNEV MQALNMSAAL TARKTREFR SPPQEQINML LNFKDDKSEC PCPEEIRDQL LDFHEDLMTH CGIELDEDGS LDGSNDLTIR GRLLSLVEKV TYLKKKQAEK PVASDSRKC SSLQQLISET MVRWAQESVI EDPELVRAMF VLLHRQYDGI GGLVRALPKT YTINGVSVED TINLLASLGQ I RSLLSVRM GKEEEKLMIR GLGDIMNNKV FYQHPNLMRA LGMHETVMEV MVNVLGGGES KEITFPKMVA NCCRFLCYFC RI SRQNQKA MFDHLSYLLE NSSVGLASPA MRGSTPLDVA AASVMDNNEL ALALREPDLE KVVRYLAGCG LQSCQMLVSK GYP DIGWNP VEGERYLDFL RFAVFCNGES VEENANVVVR LLIRRPECFG PALRGEGGNG LLAAMEEAIK IAEDPSRDGP SPTS GSSKT LDIEEEEDDT IHMGNAIMTF YAALIDLLGR CAPEMHLIHA GKGEAIRIRS ILRSLIPLGD LVGVISIAFQ MPTIA KDGK VVEPDMSAGF CPDHKAAMVL FLDRVYGIEV QDFLLHLLEV GFLPDLRAAA SLDTAALS(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)IPEKL EYFINKYAEH SHDKWSMDKL ANGWIYGEIY SDSSKIQPL MKPYKLLSEK EKEIYRWPIK ESLKTMLAWG WRIERTREGD SMALYNRTRR ISQTSQVSID AAHGYSPRAI D MSNVTLSR DLHAMAEMMA ENYHNIWAKK KKLELESKGG GNHPLLVPYD TLTAKEKAKD REKAQDIFKF LQISGYVVSR GF KDLDLDT PS(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) PRHRAVNLFL QGYEKSWIET EEHYFEDKLI EDLAK PGAE LPEEDEAMKR VDPLHQLILL FSRTALTEKC KLEEDFLYMA YADIMAKSCH DEEDDDGEEE VKSFEEKEME KQKLLY QQA RLHDRGAAEM VLQTISASKG ETGPMVAATL KLGIAILNGG NSTVQQKMLD YLKEKKDVGF FQSLAGLMQS CSVLDLN AF ERQNKAEGLG MVTEEGSGEK VLQDDEFTCD LFRFLQLLCE GHNSDFQNYL RTQTGNNTTV NIIISTVDYL LRVQESIS D FYWYYSGKDI IDEQGQRNFS KAIQVAKQVF NTLTEYIQGP CTGNQQSLAH SRLWDAVVGF LHVFAHMQMK LSQDSSQIE LLKELMDLQK DMVVMLLSML EGNVVNGTIG KQMVDMLVES SNNVEMILKF FDMFLKLKDL TSSDTFKEYD PDGKGVISKR DFHKAMESH KHYTQSETEF LLSCAETDEN ETLDYEEFVK RFHEPAKDIG FNVAVLLTNL SEHMPNDTRL QTFLELAESV L NYFQPFLG RIEIMGSAKR IERVYFEISE SSRTQWEKPQ VKESKRQFIF DVVNEGGEKE KMELFVNFCE DTIFEMQLAA QI SESDLNE RLANKEESEK ERPEEQAPRM GFFSLLTIQS ALFALRYNVL TLVRMLSLKS LKKQMKRMKK MTVKDMVLAF FSS YWSVFV TLLHFVASVC RGFFRIVSSL LLGGSLVEGA KKIKVAELLA NMPDPTQDEV RGDEEEGERK PLESALPSED LTDL KELTE ESDLLSDIFG LDLKREGGQY KLIPHNPNAG LSDLMTNPVP VPEVQEKFQE QKAKEEKEEK EETKSEPEKA EGEDG EKEE KAKDEKSKQK LRQLHTHRYG EPEVPESAFW KKIIAYQQKL LNYFARNFYN MRMLALFVAF AINFILLFYK VSTSSV VEG KELPTRTSSD TAKVTNSLDS SPHRIIAVHY VLEESSGYME PTLRILAILH TIISFFCIIG YYCLAVPLVI FKREKEV AR KLEFDGLYIT EQPSEDDIKG QWDRLVINTQ SFPNNYWDKF VKRKVMDKYG EFYGRDRISE LLGMDKAALD FSDAREKK K PKKDSSLSAV LNSIDVKYQM WKLGVVFTDN SFLYLAWYMT MSVLGHYNNF FFAAHLLDIA MGFKTLRTIL SSVTHNGKQ LVLTVGLLAV VVYLYTVVAF NFFRKFYNKS EDGDTPDMKC DDMLTCYMFH MYVGVRAGGG IGDEIEDPAG DEYEIYRIIF DITFFFFVI VILLAIIQGL IIDAFGELRD QQEQVKEDME TKCFICGIGN DYFDTVPHGF ETHTLQEHNL ANYLFFLMYL I NKDETEHT GQESYVWKMY QERCWEFFPA GDCFRKQYED QLN UniProtKB: Ryanodine receptor 2, Ryanodine receptor 2, Ryanodine receptor 2, Ryanodine receptor 2, Ryanodine receptor 2 |
-分子 #2: Peptidyl-prolyl cis-trans isomerase FKBP1B
| 分子 | 名称: Peptidyl-prolyl cis-trans isomerase FKBP1B / タイプ: protein_or_peptide / ID: 2 / コピー数: 4 / 光学異性体: LEVO / EC番号: peptidylprolyl isomerase |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 18.984316 KDa |
| 組換発現 | 生物種:  |
| 配列 | 文字列: MGSSHHHHHH SSGLVPRGSH MASMDEKTTG WRGGHVVEGL AGELEQLRAR LEHHPQGQRE PGSGGSGGTG VEIETISPGD GRTFPKKGQ TCVVHYTGML QNGKKFDSSR DRNKPFKFRI GKQEVIKGFE EGAAQMSLGQ RAKLTCTPDV AYGATGHPGV I PPNATLIF DVELLNLE UniProtKB: Peptidyl-prolyl cis-trans isomerase FKBP1B |
-分子 #3: ZINC ION
| 分子 | 名称: ZINC ION / タイプ: ligand / ID: 3 / コピー数: 4 / 式: ZN |
|---|---|
| 分子量 | 理論値: 65.409 Da |
-分子 #4: CALCIUM ION
| 分子 | 名称: CALCIUM ION / タイプ: ligand / ID: 4 / コピー数: 4 / 式: CA |
|---|---|
| 分子量 | 理論値: 40.078 Da |
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | particle |
-試料調製
| 緩衝液 | pH: 7.4 |
|---|---|
| 糖包埋 | 材質: buffer |
| 凍結 | 凍結剤: ETHANE |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | FEI TITAN KRIOS |
|---|---|
| 撮影 | フィルム・検出器のモデル: GATAN K3 (6k x 4k) / 平均電子線量: 60.0 e/Å2 |
| 電子線 | 加速電圧: 300 kV / 電子線源:  FIELD EMISSION GUN FIELD EMISSION GUN |
| 電子光学系 | 照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELD / 最大 デフォーカス(公称値): 2.0 µm / 最小 デフォーカス(公称値): 0.5 µm |
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |

