 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | 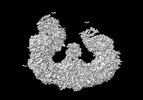 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Structure of the giant inhibitor of apoptosis, BIRC6 bound to the regulator SMAC | |||||||||
 マップデータ マップデータ | Sharpened map from non-uniform refinement | |||||||||
 試料 試料 |
| |||||||||
 キーワード キーワード | E2/E3 ubiquitin ligase / APOPTOSIS | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報activation of cysteine-type endopeptidase activity involved in apoptotic process by cytochrome c / spongiotrophoblast layer development / Release of apoptotic factors from the mitochondria / CD40 receptor complex / labyrinthine layer development / ALK mutants bind TKIs / SMAC, XIAP-regulated apoptotic response / Flemming body / Regulation of the apoptosome activity / SMAC (DIABLO) binds to IAPs ...activation of cysteine-type endopeptidase activity involved in apoptotic process by cytochrome c / spongiotrophoblast layer development / Release of apoptotic factors from the mitochondria / CD40 receptor complex / labyrinthine layer development / ALK mutants bind TKIs / SMAC, XIAP-regulated apoptotic response / Flemming body / Regulation of the apoptosome activity / SMAC (DIABLO) binds to IAPs / SMAC(DIABLO)-mediated dissociation of IAP:caspase complexes / microtubule organizing center / intrinsic apoptotic signaling pathway in response to oxidative stress / cysteine-type endopeptidase inhibitor activity / ubiquitin conjugating enzyme activity / extrinsic apoptotic signaling pathway via death domain receptors / intrinsic apoptotic signaling pathway / regulation of cytokinesis / negative regulation of extrinsic apoptotic signaling pathway / trans-Golgi network / RING-type E3 ubiquitin transferase / mitochondrial intermembrane space / cytoplasmic side of plasma membrane / spindle pole / ubiquitin-protein transferase activity / activation of cysteine-type endopeptidase activity involved in apoptotic process / Signaling by ALK fusions and activated point mutants / regulation of cell population proliferation / midbody / neuron apoptotic process / cell population proliferation / endosome / protein ubiquitination / positive regulation of apoptotic process / cell division / protein phosphorylation / centrosome / positive regulation of cell population proliferation / negative regulation of apoptotic process / apoptotic process / mitochondrion / membrane / nucleus / cytosol 類似検索 - 分子機能 | |||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.0 Å | |||||||||
 データ登録者 データ登録者 | Dietz L / Elliott PR | |||||||||
| 資金援助 |  英国, 1件 英国, 1件
| |||||||||
 引用 引用 | ジャーナル: Science / 年: 2023 タイトル: Structural basis for SMAC-mediated antagonism of caspase inhibition by the giant ubiquitin ligase BIRC6. 著者: Larissa Dietz / Cara J Ellison / Carlos Riechmann / C Keith Cassidy / F Daniel Felfoldi / Adán Pinto-Fernández / Benedikt M Kessler / Paul R Elliott / 要旨: Certain inhibitor of apoptosis (IAP) family members are sentinel proteins that prevent untimely cell death by inhibiting caspases. Antagonists, including second mitochondria-derived activator of ...Certain inhibitor of apoptosis (IAP) family members are sentinel proteins that prevent untimely cell death by inhibiting caspases. Antagonists, including second mitochondria-derived activator of caspases (SMAC), regulate IAPs and drive cell death. Baculoviral IAP repeat-containing protein 6 (BIRC6), a giant IAP with dual E2 and E3 ubiquitin ligase activity, regulates programmed cell death through unknown mechanisms. We show that BIRC6 directly restricts executioner caspase-3 and -7 and ubiquitinates caspase-3, -7, and -9, working exclusively with noncanonical E1, UBA6. Notably, we show that SMAC suppresses both mechanisms. Cryo-electron microscopy structures of BIRC6 alone and in complex with SMAC reveal that BIRC6 is an antiparallel dimer juxtaposing the substrate-binding module against the catalytic domain. Furthermore, we discover that SMAC multisite binding to BIRC6 results in a subnanomolar affinity interaction, enabling SMAC to competitively displace caspases, thus antagonizing BIRC6 anticaspase function. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
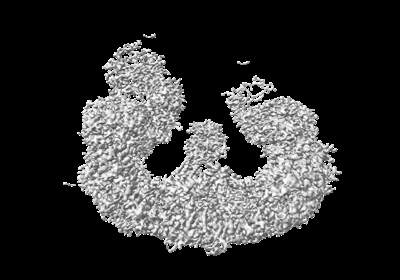
| 添付画像 |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_15654.map.gz | 95.6 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-15654-v30.xmlemd-15654.xml | 22.3 KB 22.3 KB | 表示 表示 | EMDBヘッダ |
| 画像 |  emd_15654.png emd_15654.png | 74.8 KB | ||
| マスクデータ | emd_15654_msk_1.map | 103 MB | マスクマップ | |
| Filedesc metadata | emd-15654.cif.gz | 9 KB | ||
| その他 | emd_15654_half_map_1.map.gzemd_15654_half_map_2.map.gz | 95.7 MB 95.7 MB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-15654ftp://ftp.pdbj.org/pub/emdb/structures/EMD-15654 http://ftp.pdbj.org/pub/emdb/structures/EMD-15654ftp://ftp.pdbj.org/pub/emdb/structures/EMD-15654 | HTTPS FTP |
-関連構造データ






-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| 「今月の分子」の関連する項目 |




-マップ
| ファイル | ダウンロード / ファイル: emd_15654.map.gz / 形式: CCP4 / 大きさ: 103 MB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | Sharpened map from non-uniform refinement | ||||||||||||||||||||
| ボクセルのサイズ | X=Y=Z: 0.829 Å | ||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||
| 詳細 | EMDB XML:
|
-添付データ
-マスク #1
| ファイル | emd_15654_msk_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 |
| ||||||||||||
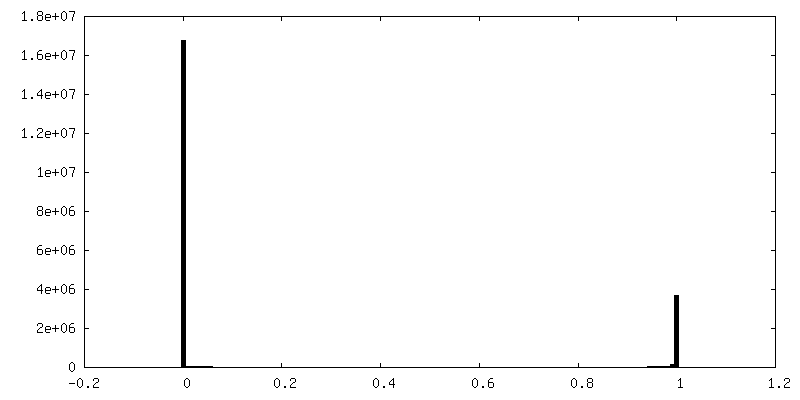
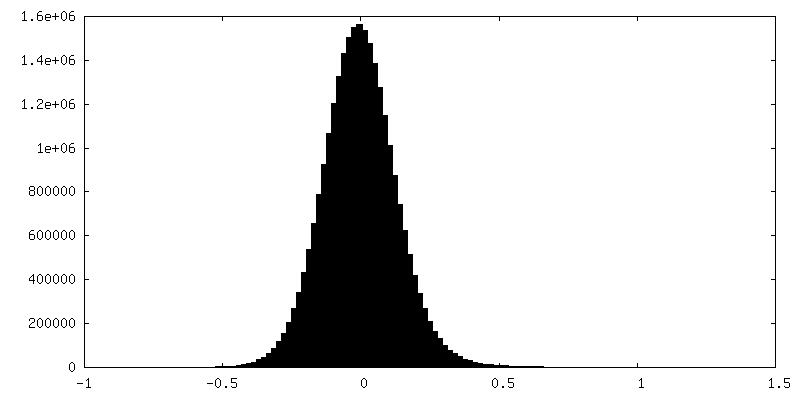
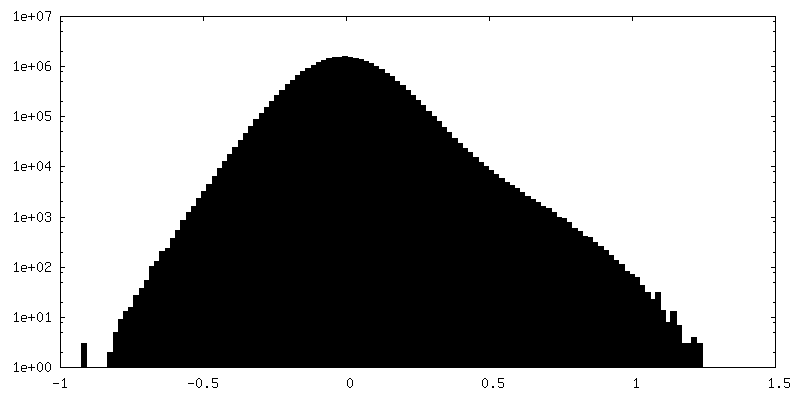
| 密度ヒストグラム |
 Z
Z Y
Y X
X





-ハーフマップ: First half map non-uniform refinement
| ファイル | emd_15654_half_map_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | First half map non-uniform refinement | ||||||||||||
| 投影像・断面図 |
| ||||||||||||
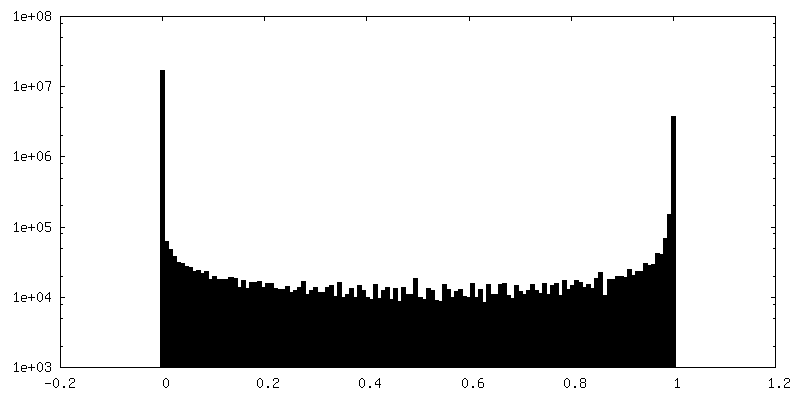
| 密度ヒストグラム |






-ハーフマップ: Second half map non-uniform refinement
| ファイル | emd_15654_half_map_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | Second half map non-uniform refinement | ||||||||||||
| 投影像・断面図 |
| ||||||||||||
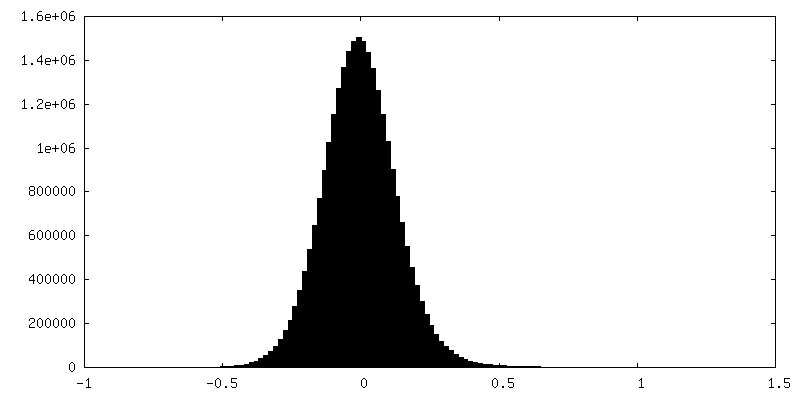
| 密度ヒストグラム |

- 試料の構成要素
試料の構成要素
-全体 : Anti-parallel homodimer
| 全体 | 名称: Anti-parallel homodimer |
|---|---|
| 要素 |
|
-超分子 #1: Anti-parallel homodimer
| 超分子 | 名称: Anti-parallel homodimer / タイプ: complex / ID: 1 / 親要素: 0 / 含まれる分子: all |
|---|---|
| 分子量 | 理論値: 1 MDa |
-超分子 #2: Baculoviral IAP repeat-containing protein 6
| 超分子 | 名称: Baculoviral IAP repeat-containing protein 6 / タイプ: complex / ID: 2 / 親要素: 1 / 含まれる分子: #1 |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
-超分子 #3: Diablo IAP-binding mitochondrial protein
| 超分子 | 名称: Diablo IAP-binding mitochondrial protein / タイプ: complex / ID: 3 / 親要素: 1 / 含まれる分子: #2 |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
-分子 #1: Baculoviral IAP repeat-containing protein 6
| 分子 | 名称: Baculoviral IAP repeat-containing protein 6 / タイプ: protein_or_peptide / ID: 1 / コピー数: 2 / 光学異性体: LEVO / EC番号: RING-type E3 ubiquitin transferase |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 530.977 KDa |
| 組換発現 | 生物種:   Spodoptera frugiperda (ツマジロクサヨトウ) Spodoptera frugiperda (ツマジロクサヨトウ) |
| 配列 | 文字列: GPMVTGGGAA PPGTVTEPLP SVIVLSAGRK MAAAAAAASG PGCSSAAGAG AAGVSEWLVL RDGCMHCDAD GLHSLSYHPA LNAILAVTS RGTIKVIDGT SGATLQASAL SAKPGGQVKC QYISAVDKVI FVDDYAVGCR KDLNGILLLD TALQTPVSKQ D DVVQLELP ...文字列: GPMVTGGGAA PPGTVTEPLP SVIVLSAGRK MAAAAAAASG PGCSSAAGAG AAGVSEWLVL RDGCMHCDAD GLHSLSYHPA LNAILAVTS RGTIKVIDGT SGATLQASAL SAKPGGQVKC QYISAVDKVI FVDDYAVGCR KDLNGILLLD TALQTPVSKQ D DVVQLELP VTEAQQLLSA CLEKVDISST EGYDLFITQL KDGLKNTSHE TAANHKVAKW ATVTFHLPHH VLKSIASAIV NE LKKINQN VAALPVASSV MDRLSYLLPS ARPELGVGPG RSVDRSLMYS EANRRETFTS WPHVGYRWAQ PDPMAQAGFY HQP ASSGDD RAMCFTCSVC LVCWEPTDEP WSEHERHSPN CPFVKGEHTQ NVPLSVTLAT SPAQFPCTDG TDRISCFGSG SCPH FLAAA TKRGKICIWD VSKLMKVHLK FEINAYDPAI VQQLILSGDP SSGVDSRRPT LAWLEDSSSC SDIPKLEGDS DDLLE DSDS EEHSRSDSVT GHTSQKEAME VSLDITALSI LQQPEKLQWE IVANVLEDTV KDLEELGANP CLTNSKSEKT KEKHQE QHN IPFPCLLAGG LLTYKSPATS PISSNSHRSL DGLSRTQGES ISEQGSTDNE SCTNSELNSP LVRRTLPVLL LYSIKES DE KAGKIFSQMN NIMSKSLHDD GFTVPQIIEM ELDSQEQLLL QDPPVTYIQQ FADAAANLTS PDSEKWNSVF PKPGTLVQ C LRLPKFAEEE NLCIDSITPC ADGIHLLVGL RTCPVESLSA INQVEALNNL NKLNSALCNR RKGELESNLA VVNGANISV IQHESPADVQ TPLIIQPEQR NVSGGYLVLY KMNYATRIVT LEEEPIKIQH IKDPQDTITS LILLPPDILD NREDDCEEPI EDMQLTSKN GFEREKTSDI STLGHLVITT QGGYVKILDL SNFEILAKVE PPKKEGTEEQ DTFVSVIYCS GTDRLCACTK G GELHFLQI GGTCDDIDEA DILVDGSLSK GIEPSSEGSK PLSNPSSPGI SGVDLLVDQP FTLEILTSLV ELTRFETLTP RF SATVPPC WVEVQQEQQQ RRHPQHLHQQ HHGDAAQHTR TWKLQTDSNS WDEHVFELVL PKACMVGHVD FKFVLNSNIT NIP QIQVTL LKNKAPGLGK VNALNIEVEQ NGKPSLVDLN EEMQHMDVEE SQCLRLCPFL EDHKEDILCG PVWLASGLDL SGHA GMLTL TSPKLVKGMA GGKYRSFLIH VKAVNERGTE EICNGGMRPV VRLPSLKHQS NKGYSLASLL AKVAAGKEKS SNVKN ENTS GTRKSENLRG CDLLQEVSVT IRRFKKTSIS KERVQRCAML QFSEFHEKLL NTLCRKTDDG QITEHAQSLV LDTLCW LAG VHSNGPGSSK EGNENLLSKT RKFLSDIVRV CFFEAGRSIA HKCARFLALC ISNGKCDPCQ PAFGPVLLKA LLDNMSF LP AATTGGSVYW YFVLLNYVKD EDLAGCSTAC ASLLTAVSRQ LQDRLTPMEA LLQTRYGLYS SPFDPVLFDL EMSGSSCK N VYNSSIGVQS DEIDLSDVLS GNGKVSSCTA AEGSFTSLTG LLEVEPLHFT CVSTSDGTRI ERDDAMSSFG VTPAVGGLS SGTVGEASTA LSSAAQVALQ SLSHAMASAE QQLQVLQEKQ QQLLKLQQQK AKLEAKLHQT TAAAAAAASA VGPVHNSVPS NPVAAPGFF IHPSDVIPPT PKTTPLFMTP PLTPPNEAVS VVINAELAQL FPGSVIDPPA VNLAAHNKNS NKSRMNPLGS G LALAISHA SHFLQPPPHQ SIIIERMHSG ARRFVTLDFG RPILLTDVLI PTCGDLASLS IDIWTLGEEV DGRRLVVATD IS THSLILH DLIPPPVCRF MKITVIGRYG STNARAKIPL GFYYGHTYIL PWESELKLMH DPLKGEGESA NQPEIDQHLA MMV ALQEDI QCRYNLACHR LETLLQSIDL PPLNSANNAQ YFLRKPDKAV EEDSRVFSAY QDCIQLQLQL NLAHNAVQRL KVAL GASRK MLSETSNPED LIQTSSTEQL RTIIRYLLDT LLSLLHASNG HSVPAVLQST FHAQACEELF KHLCISGTPK IRLHT GLLL VQLCGGERWW GQFLSNVLQE LYNSEQLLIF PQDRVFMLLS CIGQRSLSNS GVLESLLNLL DNLLSPLQPQ LPMHRR TEG VLDIPMISWV VMLVSRLLDY VATVEDEAAA AKKPLNGNQW SFINNNLHTQ SLNRSSKGSS SLDRLYSRKI RKQLVHH KQ QLNLLKAKQK ALVEQMEKEK IQSNKGSSYK LLVEQAKLKQ ATSKHFKDLI RLRRTAEWSR SNLDTEVTTA KESPEIEP L PFTLAHERCI SVVQKLVLFL LSMDFTCHAD LLLFVCKVLA RIANATRPTI HLCEIVNEPQ LERLLLLLVG TDFNRGDIS WGGAWAQYSL TCMLQDILAG ELLAPVAAEA MEEGTVGDDV GATAGDSDDS LQQSSVQLLE TIDEPLTHDI TGAPPLSSLE KDKEIDLEL LQDLMEVDID PLDIDLEKDP LAAKVFKPIS STWYDYWGAD YGTYNYNPYI GGLGIPVAKP PANTEKNGSQ T VSVSVSQA LDARLEVGLE QQAELMLKMM STLEADSILQ ALTNTSPTLS QSPTGTDDSL LGGLQAANQT SQLIIQLSSV PM LNVCFNK LFSMLQVHHV QLESLLQLWL TLSLNSSSTG NKENGADIFL YNANRIPVIS LNQASITSFL TVLAWYPNTL LRT WCLVLH SLTLMTNMQL NSGSSSAIGT QESTAHLLVS DPNLIHVLVK FLSGTSPHGT NQHSPQVGPT ATQAMQEFLT RLQV HLSST CPQIFSEFLL KLIHILSTER GAFQTGQGPL DAQVKLLEFT LEQNFEVVSV STISAVIESV TFLVHHYITC SDKVM SRSG SDSSVGARAC FGGLFANLIR PGDAKAVCGE MTRDQLMFDL LKLVNILVQL PLSGNREYSA RVSVTTNTTD SVSDEE KVS GGKDGNGSST SVQGSPAYVA DLVLANQQIM SQILSALGLC NSSAMAMIIG ASGLHLTKHE NFHGGLDAIS VGDGLFT IL TTLSKKASTV HMMLQPILTY MACGYMGRQG SLATCQLSEP LLWFILRVLD TSDALKAFHD MGGVQLICNN MVTSTRAI V NTARSMVSTI MKFLDSGPNK AVDSTLKTRI LASEPDNAEG IHNFAPLGTI TSSSPTAQPA EVLLQATPPH RRARSAAWS YIFLPEEAWC DLTIHLPAAV LLKEIHIQPH LASLATCPSS VSVEVSADGV NMLPLSTPVV TSGLTYIKIQ LVKAEVASAV CLRLHRPRD ASTLGLSQIK LLGLTAFGTT SSATVNNPFL PSEDQVSKTS IGWLRLLHHC LTHISDLEGM MASAAAPTAN L LQTCAALL MSPYCGMHSP NIEVVLVKIG LQSTRIGLKL IDILLRNCAA SGSDPTDLNS PLLFGRLNGL SSDSTIDILY QL GTTQDPG TKDRIQALLK WVSDSARVAA MKRSGRMNYM CPNSSTVEYG LLMPSPSHLH CVAAILWHSY ELLVEYDLPA LLD QELFEL LFNWSMSLPC NMVLKKAVDS LLCSMCHVHP NYFSLLMGWM GITPPPVQCH HRLSMTDDSK KQDLSSSLTD DSKN AQAPL ALTESHLATL ASSSQSPEAI KQLLDSGLPS LLVRSLASFC FSHISSSESI AQSIDISQDK LRRHHVPQQC NKMPI TADL VAPILRFLTE VGNSHIMKDW LGGSEVNPLW TALLFLLCHS GSTSGSHNLG AQQTSARSAS LSSAATTGLT TQQRTA IEN ATVAFFLQCI SCHPNNQKLM AQVLCELFQT SPQRGNLPTS GNISGFIRRL FLQLMLEDEK VTMFLQSPCP LYKGRIN AT SHVIQHPMYG AGHKFRTLHL PVSTTLSDVL DRVSDTPSIT AKLISEQKDD KEKKNHEEKE KVKAENGFQD NYSVVVAS G LKSQSKRAVS ATPPRPPSRR GRTIPDKIGS TSGAEAANKI ITVPVFHLFH KLLAGQPLPA EMTLAQLLTL LYDRKLPQG YRSIDLTVKL GSRVITDPSL SKTDSYKRLH PEKDHGDLLA SCPEDEALTP GDECMDGILD ESLLETCPIQ SPLQVFAGMG GLALIAERL PMLYPEVIQQ VSAPVVTSTT QEKPKDSDQF EWVTIEQSGE LVYEAPETVA AEPPPIKSAV QTMSPIPAHS L AAFGLFLR LPGYAEVLLK ERKHAQCLLR LVLGVTDDGE GSHILQSPSA NVLPTLPFHV LRSLFSTTPL TTDDGVLLRR MA LEIGALH LILVCLSALS HHSPRVPNSS VNQTEPQVSS SHNPTSTEEQ QLYWAKGTGF GTGSTASGWD VEQALTKQRL EEE HVTCLL QVLASYINPV SSAVNGEAQS SHETRGQNSN ALPSVLLELL SQSCLIPAMS SYLRNDSVLD MARHVPLYRA LLEL LRAIA SCAAMVPLLL PLSTENGEEE EEQSECQTSV GTLLAKMKTC VDTYTNRLRS KRENVKTGVK PDASDQEPEG LTLLV PDIQ KTAEIVYAAT TSLRQANQEK KLGEYSKKAA MKPKPLSVLK SLEEKYVAVM KKLQFDTFEM VSEDEDGKLG FKVNYH YMS QVKNANDANS AARARRLAQE AVTLSTSLPL SSSSSVFVRC DEERLDIMKV LITGPADTPY ANGCFEFDVY FPQDYPS SP PLVNLETTGG HSVRFNPNLY NDGKVCLSIL NTWHGRPEEK WNPQTSSFLQ VLVSVQSLIL VAEPYFNEPG YERSRGTP S GTQSSREYDG NIRQATVKWA MLEQIRNPSP CFKEVIHKHF YLKRVEIMAQ CEEWIADIQQ YSSDKRVGRT MSHHAAALK RHTAQLREEL LKLPCPEGLD PDTDDAPEVC RATTGAEETL MHDQVKPSSS KELPSDFQL UniProtKB: Baculoviral IAP repeat-containing protein 6 |
-分子 #2: Diablo IAP-binding mitochondrial protein
| 分子 | 名称: Diablo IAP-binding mitochondrial protein / タイプ: protein_or_peptide / ID: 2 / コピー数: 2 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 20.787098 KDa |
| 組換発現 | 生物種:  |
| 配列 | 文字列: AVPIAQKSEP HSLSSEALMR RAVSLVTDST STFLSQTTYA LIEAITEYTK AVYTLTSLYR QYTSLLGKMN SEEEDEVWQV IIGARAEMT SKHQEYLKLE TTWMTAVGLS EMAAEAAYQT GADQASITAR NHIQLVKLQV EEVHQLSRKA ETKLAEAQIE E LRQKTQEE GEERAESEQE AYLRED UniProtKB: Diablo IAP-binding mitochondrial protein |
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | particle |
-試料調製
| 濃度 | 4 mg/mL |
|---|---|
| 緩衝液 | pH: 7.5 |
| 凍結 | 凍結剤: ETHANE / チャンバー内湿度: 100 % / チャンバー内温度: 277 K / 装置: FEI VITROBOT MARK III |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | FEI TITAN KRIOS |
|---|---|
| 撮影 | フィルム・検出器のモデル: GATAN K3 (6k x 4k) / 平均露光時間: 2.3 sec. / 平均電子線量: 47.27 e/Å2 |
| 電子線 | 加速電圧: 300 kV / 電子線源:  FIELD EMISSION GUN FIELD EMISSION GUN |
| 電子光学系 | C2レンズ絞り径: 70.0 µm / 最大 デフォーカス(補正後): 2.5 µm / 最小 デフォーカス(補正後): 0.75 µm / 照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELD / Cs: 2.7 mm / 最大 デフォーカス(公称値): 2.5 µm / 最小 デフォーカス(公称値): 0.75 µm / 倍率(公称値): 105000 |
| 試料ステージ | 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER ホルダー冷却材: NITROGEN |
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
+画像解析
-原子モデル構築 1
| 精密化 | 空間: REAL / プロトコル: OTHER / 温度因子: 191.06 |
|---|---|
| 得られたモデル |  PDB-8ato: |
