National Institutes of Health/National Cancer Institute (NIH/NCI)
U54 CA209891
米国
National Institutes of Health/National Institute of General Medical Sciences (NIH/NIGMS)
R35 GM139636
米国
引用
ジャーナル: Nat Commun / 年: 2023 タイトル: Structural insights into regulation of the PEAK3 pseudokinase scaffold by 14-3-3. 著者: Hayarpi Torosyan / Michael D Paul / Antoine Forget / Megan Lo / Devan Diwanji / Krzysztof Pawłowski / Nevan J Krogan / Natalia Jura / Kliment A Verba / 要旨: PEAK pseudokinases are molecular scaffolds which dimerize to regulate cell migration, morphology, and proliferation, as well as cancer progression. The mechanistic role dimerization plays in PEAK ...PEAK pseudokinases are molecular scaffolds which dimerize to regulate cell migration, morphology, and proliferation, as well as cancer progression. The mechanistic role dimerization plays in PEAK scaffolding remains unclear, as there are no structures of PEAKs in complex with their interactors. Here, we report the cryo-EM structure of dimeric PEAK3 in complex with an endogenous 14-3-3 heterodimer. Our structure reveals an asymmetric binding mode between PEAK3 and 14-3-3 stabilized by one pseudokinase domain and the SHED domain of the PEAK3 dimer. The binding interface contains a canonical phosphosite-dependent primary interaction and a unique secondary interaction not observed in previous structures of 14-3-3/client complexes. Additionally, we show that PKD regulates PEAK3/14-3-3 binding, which when prevented leads to PEAK3 nuclear enrichment and distinct protein-protein interactions. Altogether, our data demonstrate that PEAK3 dimerization forms an unusual secondary interface for 14-3-3 binding, facilitating 14-3-3 regulation of PEAK3 localization and interactome diversity.
14-3-3proteinbeta/alpha / Protein 1054 / Protein kinase C inhibitor protein 1 / KCIP-1
分子量: 28114.373 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: YWHAB / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: P31946
#3: タンパク質
14-3-3proteinepsilon / 14-3-3E
分子量: 29208.900 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: YWHAE / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: P62258
#4: タンパク質
ProteinPEAK3fragment
分子量: 52437.012 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: PEAK3, C19orf35 / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: Q6ZS72
構成要素の詳細
The authors state that chains E and P are part of Chains A and B. However, because they cannot ...The authors state that chains E and P are part of Chains A and B. However, because they cannot resolve a large portion of the N-terminal segments of Chain A and B and therefore the connectivity between these two sets of chains, they cannot with confidence assign Chain E residues to Chains A or B and the same with Chain P residues.
研究の焦点であるリガンドがあるか
N
Has protein modification
Y
-
実験情報
-
実験
実験
手法: 電子顕微鏡法
EM実験
試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法
-
試料調製
構成要素
名称: Complex between PEAK3 and 14-3-3 epsilon, beta / タイプ: COMPLEX / Entity ID: all / 由来: RECOMBINANT
分子量
値: 0.1618 MDa / 実験値: NO
由来(天然)
生物種: Homo sapiens (ヒト)
由来(組換発現)
生物種: Homo sapiens (ヒト)
緩衝液
pH: 7.5 詳細: A final concentration of 0.1% of Octyl-beta-Glucoside (C14H28O6) was added to the sample before freezing.
緩衝液成分
ID
濃度
名称
式
Buffer-ID
1
50mM
TrisBase
NH2C(CH2OH)3
1
2
150mM
SodiumChloride
NaCl
1
3
2mM
Dithiothreitol
C4H10O2S2
1
4
2mM
MagnesiumChloride
MgCl2
1
試料
濃度: 1.1 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) データ登録者
データ登録者 米国, 3件
米国, 3件  引用
引用
 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード


 PDBj
PDBj











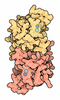



 集合体
集合体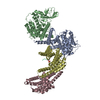
 試料調製
試料調製 電子顕微鏡撮影
電子顕微鏡撮影
 FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM
FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM 解析
解析