 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8dp5 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Structure of the PEAK3/14-3-3 complex | ||||||||||||
 Components Components |
| ||||||||||||
 Keywords Keywords | SIGNALING PROTEIN / complex / pseudokinase / kinase / adapter | ||||||||||||
| Function / homology |  Function and homology information Function and homology informationregulation of heart rate by hormone / Tristetraprolin (TTP, ZFP36) binds and destabilizes mRNA / positive regulation of hippo signaling / Butyrate Response Factor 1 (BRF1) binds and destabilizes mRNA / membrane repolarization during cardiac muscle cell action potential / negative regulation of toll-like receptor signaling pathway / regulation of membrane repolarization / NADE modulates death signalling / protein localization to endoplasmic reticulum / regulation of cell motility ...regulation of heart rate by hormone / Tristetraprolin (TTP, ZFP36) binds and destabilizes mRNA / positive regulation of hippo signaling / Butyrate Response Factor 1 (BRF1) binds and destabilizes mRNA / membrane repolarization during cardiac muscle cell action potential / negative regulation of toll-like receptor signaling pathway / regulation of membrane repolarization / NADE modulates death signalling / protein localization to endoplasmic reticulum / regulation of cell motility / MTOR signalling / regulation of potassium ion transmembrane transport / ARMS-mediated activation / RAB GEFs exchange GTP for GDP on RABs / SHOC2 M1731 mutant abolishes MRAS complex function / Gain-of-function MRAS complexes activate RAF signaling / Rap1 signalling / Signaling by Hippo / vacuolar membrane / negative regulation of protein import into nucleus / negative regulation of G protein-coupled receptor signaling pathway / Frs2-mediated activation / protein phosphatase inhibitor activity / Deregulated CDK5 triggers multiple neurodegenerative pathways in Alzheimer's disease models / negative regulation of calcium ion export across plasma membrane / cytoplasmic pattern recognition receptor signaling pathway / intracellular potassium ion homeostasis / protein kinase inhibitor activity / Regulation of localization of FOXO transcription factors / regulation of heart rate by cardiac conduction / mTORC1-mediated signalling / phosphoserine residue binding / Activation of BAD and translocation to mitochondria / Regulation of HSF1-mediated heat shock response / potassium channel regulator activity / HSF1 activation / protein localization to nucleus / SARS-CoV-2 targets host intracellular signalling and regulatory pathways / regulation of cytosolic calcium ion concentration / calcium channel inhibitor activity / Chk1/Chk2(Cds1) mediated inactivation of Cyclin B:Cdk1 complex / SARS-CoV-1 targets host intracellular signalling and regulatory pathways / RHO GTPases activate PKNs / transcription repressor complex / Loss of Nlp from mitotic centrosomes / Loss of proteins required for interphase microtubule organization from the centrosome / Recruitment of mitotic centrosome proteins and complexes / Transcriptional and post-translational regulation of MITF-M expression and activity / Recruitment of NuMA to mitotic centrosomes / substantia nigra development / Anchoring of the basal body to the plasma membrane / signaling adaptor activity / regulation of mitotic cell cycle / positive regulation of protein export from nucleus / AURKA Activation by TPX2 / regulation of actin cytoskeleton organization / TP53 Regulates Metabolic Genes / Translocation of SLC2A4 (GLUT4) to the plasma membrane / calcium channel regulator activity / protein sequestering activity / phosphoprotein binding / RAF activation / Signaling by high-kinase activity BRAF mutants / MAP2K and MAPK activation / mitochondrial membrane / histone deacetylase binding / regulation of cell shape / intracellular protein localization / Signaling by RAF1 mutants / Signaling by moderate kinase activity BRAF mutants / Paradoxical activation of RAF signaling by kinase inactive BRAF / Signaling downstream of RAS mutants / MAPK cascade / Negative regulation of MAPK pathway / melanosome / Signaling by BRAF and RAF1 fusions / MHC class II protein complex binding / actin cytoskeleton / Regulation of PLK1 Activity at G2/M Transition / cellular response to heat / scaffold protein binding / protein phosphatase binding / transmembrane transporter binding / protein kinase activity / intracellular signal transduction / cadherin binding / protein heterodimerization activity / protein domain specific binding / focal adhesion / negative regulation of DNA-templated transcription / ubiquitin protein ligase binding / protein-containing complex binding / perinuclear region of cytoplasm / enzyme binding / signal transduction / endoplasmic reticulum / positive regulation of transcription by RNA polymerase II / RNA binding / extracellular exosome / membrane Similarity search - Function | ||||||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.1 Å | ||||||||||||
 Authors Authors | Torosyan, H. / Paul, M. / Jura, N. / Verba, K.A. | ||||||||||||
| Funding support |  United States, 3items United States, 3items
| ||||||||||||
 Citation Citation | Journal: Nat Commun / Year: 2023 Title: Structural insights into regulation of the PEAK3 pseudokinase scaffold by 14-3-3. Authors: Hayarpi Torosyan / Michael D Paul / Antoine Forget / Megan Lo / Devan Diwanji / Krzysztof Pawłowski / Nevan J Krogan / Natalia Jura / Kliment A Verba /  Abstract: PEAK pseudokinases are molecular scaffolds which dimerize to regulate cell migration, morphology, and proliferation, as well as cancer progression. The mechanistic role dimerization plays in PEAK ...PEAK pseudokinases are molecular scaffolds which dimerize to regulate cell migration, morphology, and proliferation, as well as cancer progression. The mechanistic role dimerization plays in PEAK scaffolding remains unclear, as there are no structures of PEAKs in complex with their interactors. Here, we report the cryo-EM structure of dimeric PEAK3 in complex with an endogenous 14-3-3 heterodimer. Our structure reveals an asymmetric binding mode between PEAK3 and 14-3-3 stabilized by one pseudokinase domain and the SHED domain of the PEAK3 dimer. The binding interface contains a canonical phosphosite-dependent primary interaction and a unique secondary interaction not observed in previous structures of 14-3-3/client complexes. Additionally, we show that PKD regulates PEAK3/14-3-3 binding, which when prevented leads to PEAK3 nuclear enrichment and distinct protein-protein interactions. Altogether, our data demonstrate that PEAK3 dimerization forms an unusual secondary interface for 14-3-3 binding, facilitating 14-3-3 regulation of PEAK3 localization and interactome diversity. | ||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8dp5.cif.gz | 435.5 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8dp5.ent.gz | 351.2 KB | Display | PDB format |
| PDBx/mmJSON format | 8dp5.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/dp/8dp5ftp://data.pdbj.org/pub/pdb/validation_reports/dp/8dp5 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  27630MC  8ds6C M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |

-Links
 PDBj
PDBj











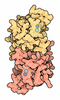



- Assembly
Assembly
| Deposited unit | 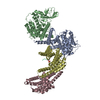
|
|---|---|
| 1 |
|
-Components
| #1: Protein | Mass: 52357.031 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: PEAK3, C19orf35 / Cell line (production host): Expi293 / Production host: Homo sapiens (human) / References: UniProt: Q6ZS72#2: Protein | | Mass: 28114.373 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: YWHAB / Production host: Homo sapiens (human) / References: UniProt: P31946#3: Protein | | Mass: 29208.900 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: YWHAE / Production host: Homo sapiens (human) / References: UniProt: P62258#4: Protein | Mass: 52437.012 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: PEAK3, C19orf35 / Production host: Homo sapiens (human) / References: UniProt: Q6ZS72Compound details | The authors state that chains E and P are part of Chains A and B. However, because they cannot ...The authors state that chains E and P are part of Chains A and B. However, because they cannot resolve a large portion of the N-terminal segments of Chain A and B and therefore the connectivity between these two sets of chains, they cannot with confidence assign Chain E residues to Chains A or B and the same with Chain P residues. | Has ligand of interest | N | Has protein modification | Y | |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: Complex between PEAK3 and 14-3-3 epsilon, beta / Type: COMPLEX / Entity ID: all / Source: RECOMBINANT | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Value: 0.1618 MDa / Experimental value: NO | |||||||||||||||||||||||||
| Source (natural) | Organism: Homo sapiens (human) | |||||||||||||||||||||||||
| Source (recombinant) | Organism: Homo sapiens (human) | |||||||||||||||||||||||||
| Buffer solution | pH: 7.5 Details: A final concentration of 0.1% of Octyl-beta-Glucoside (C14H28O6) was added to the sample before freezing. | |||||||||||||||||||||||||
| Buffer component |
| |||||||||||||||||||||||||
| Specimen | Conc.: 1.1 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | |||||||||||||||||||||||||
| Specimen support | Grid material: GOLD / Grid mesh size: 300 divisions/in. / Grid type: Quantifoil R1.2/1.3 | |||||||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK IV / Cryogen name: ETHANE / Humidity: 100 % / Chamber temperature: 278.15 K / Details: blot time = 7s blot force = 4 |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 105000 X / Nominal defocus max: 2000 nm / Nominal defocus min: 1000 nm / Alignment procedure: COMA FREE |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Electron dose: 69 e/Å2 / Film or detector model: GATAN K3 (6k x 4k) / Num. of grids imaged: 1 |
| EM imaging optics | Energyfilter name: GIF Bioquantum / Energyfilter slit width: 20 eV |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.1 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 169563 / Symmetry type: POINT |
