 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7zu0 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
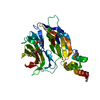
| タイトル | HOPS tethering complex from yeast | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | CYTOSOLIC PROTEIN / HOPS / tethering complex / lysosome / membrane fusion / Rab GTPase / cryo-EM | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報histone catabolic process / CORVET complex / organelle fusion / HOPS complex / vesicle tethering / endosomal vesicle fusion / regulation of vacuole fusion, non-autophagic / vacuole-mitochondrion membrane contact site / vacuolar protein processing / regulation of SNARE complex assembly ...histone catabolic process / CORVET complex / organelle fusion / HOPS complex / vesicle tethering / endosomal vesicle fusion / regulation of vacuole fusion, non-autophagic / vacuole-mitochondrion membrane contact site / vacuolar protein processing / regulation of SNARE complex assembly / vesicle fusion with vacuole / vacuole fusion, non-autophagic / cytoplasm to vacuole targeting by the Cvt pathway / Golgi to vacuole transport / Golgi to endosome transport / vesicle docking / vacuole organization / protein targeting to vacuole / late endosome to vacuole transport / fungal-type vacuole / endosome organization / piecemeal microautophagy of the nucleus / vesicle docking involved in exocytosis / fungal-type vacuole membrane / endosomal transport / vesicle-mediated transport / positive regulation of TORC1 signaling / guanyl-nucleotide exchange factor activity / cellular response to starvation / macroautophagy / intracellular protein transport / RING-type E3 ubiquitin transferase / autophagy / small GTPase binding / endocytosis / ubiquitin protein ligase activity / late endosome / protein transport / actin binding / early endosome membrane / protein-macromolecule adaptor activity / endosome / zinc ion binding / ATP binding / cytoplasm / cytosol 類似検索 - 分子機能 | ||||||||||||
| 生物種 |  | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 4.4 Å | ||||||||||||
 データ登録者 データ登録者 | Shvarev, D. / Schoppe, J. / Koenig, C. / Perz, A. / Fuellbrunn, N. / Kiontke, S. / Langemeyer, L. / Januliene, D. / Schnelle, K. / Kuemmel, D. ...Shvarev, D. / Schoppe, J. / Koenig, C. / Perz, A. / Fuellbrunn, N. / Kiontke, S. / Langemeyer, L. / Januliene, D. / Schnelle, K. / Kuemmel, D. / Froehlich, F. / Moeller, A. / Ungermann, C. | ||||||||||||
| 資金援助 |  ドイツ, 3件 ドイツ, 3件
| ||||||||||||
 引用 引用 | ジャーナル: Elife / 年: 2022 タイトル: Structure of the HOPS tethering complex, a lysosomal membrane fusion machinery. 著者: Dmitry Shvarev / Jannis Schoppe / Caroline König / Angela Perz / Nadia Füllbrunn / Stephan Kiontke / Lars Langemeyer / Dovile Januliene / Kilian Schnelle / Daniel Kümmel / Florian ...著者: Dmitry Shvarev / Jannis Schoppe / Caroline König / Angela Perz / Nadia Füllbrunn / Stephan Kiontke / Lars Langemeyer / Dovile Januliene / Kilian Schnelle / Daniel Kümmel / Florian Fröhlich / Arne Moeller / Christian Ungermann / 要旨: Lysosomes are essential for cellular recycling, nutrient signaling, autophagy, and pathogenic bacteria and viruses invasion. Lysosomal fusion is fundamental to cell survival and requires HOPS, a ...Lysosomes are essential for cellular recycling, nutrient signaling, autophagy, and pathogenic bacteria and viruses invasion. Lysosomal fusion is fundamental to cell survival and requires HOPS, a conserved heterohexameric tethering complex. On the membranes to be fused, HOPS binds small membrane-associated GTPases and assembles SNAREs for fusion, but how the complex fulfills its function remained speculative. Here, we used cryo-electron microscopy to reveal the structure of HOPS. Unlike previously reported, significant flexibility of HOPS is confined to its extremities, where GTPase binding occurs. The SNARE-binding module is firmly attached to the core, therefore, ideally positioned between the membranes to catalyze fusion. Our data suggest a model for how HOPS fulfills its dual functionality of tethering and fusion and indicate why it is an essential part of the membrane fusion machinery. #1: ジャーナル: Nat Commun / 年: 2024タイトル: Structure of the endosomal CORVET tethering complex. 著者: Dmitry Shvarev / Caroline König / Nicole Susan / Lars Langemeyer / Stefan Walter / Angela Perz / Florian Fröhlich / Christian Ungermann / Arne Moeller / 要旨: Cells depend on their endolysosomal system for nutrient uptake and downregulation of plasma membrane proteins. These processes rely on endosomal maturation, which requires multiple membrane fusion ...Cells depend on their endolysosomal system for nutrient uptake and downregulation of plasma membrane proteins. These processes rely on endosomal maturation, which requires multiple membrane fusion steps. Early endosome fusion is promoted by the Rab5 GTPase and its effector, the hexameric CORVET tethering complex, which is homologous to the lysosomal HOPS. How these related complexes recognize their specific target membranes remains entirely elusive. Here, we solve the structure of CORVET by cryo-electron microscopy and revealed its minimal requirements for membrane tethering. As expected, the core of CORVET and HOPS resembles each other. However, the function-defining subunits show marked structural differences. Notably, we discover that unlike HOPS, CORVET depends not only on Rab5 but also on phosphatidylinositol-3-phosphate (PI3P) and membrane lipid packing defects for tethering, implying that an organelle-specific membrane code enables fusion. Our data suggest that both shape and membrane interactions of CORVET and HOPS are conserved in metazoans, thus providing a paradigm how tethering complexes function. #2: ジャーナル: Elife / 年: 2022タイトル: Structure of the HOPS tethering complex, a lysosomal membrane fusion machinery. 著者: Dmitry Shvarev / Jannis Schoppe / Caroline König / Angela Perz / Nadia Füllbrunn / Stephan Kiontke / Lars Langemeyer / Dovile Januliene / Kilian Schnelle / Daniel Kümmel / Florian ...著者: Dmitry Shvarev / Jannis Schoppe / Caroline König / Angela Perz / Nadia Füllbrunn / Stephan Kiontke / Lars Langemeyer / Dovile Januliene / Kilian Schnelle / Daniel Kümmel / Florian Fröhlich / Arne Moeller / Christian Ungermann / 要旨: Lysosomes are essential for cellular recycling, nutrient signaling, autophagy, and pathogenic bacteria and viruses invasion. Lysosomal fusion is fundamental to cell survival and requires HOPS, a ...Lysosomes are essential for cellular recycling, nutrient signaling, autophagy, and pathogenic bacteria and viruses invasion. Lysosomal fusion is fundamental to cell survival and requires HOPS, a conserved heterohexameric tethering complex. On the membranes to be fused, HOPS binds small membrane-associated GTPases and assembles SNAREs for fusion, but how the complex fulfills its function remained speculative. Here, we used cryo-electron microscopy to reveal the structure of HOPS. Unlike previously reported, significant flexibility of HOPS is confined to its extremities, where GTPase binding occurs. The SNARE-binding module is firmly attached to the core, therefore, ideally positioned between the membranes to catalyze fusion. Our data suggest a model for how HOPS fulfills its dual functionality of tethering and fusion and indicate why it is an essential part of the membrane fusion machinery. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7zu0.cif.gz | 649.7 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7zu0.ent.gz | 484.2 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 7zu0.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/zu/7zu0ftp://data.pdbj.org/pub/pdb/validation_reports/zu/7zu0 | HTTPS FTP |
|---|
-関連構造データ







-リンク
 PDBj
PDBj



- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 3種, 3分子 ACE
| #1: タンパク質 | 分子量: 117617.219 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c 遺伝子: PEP5, END1, VAM1, VPL9, VPS11, VPT11, YMR231W, YM9959.13 発現宿主: |
|---|---|
| #3: タンパク質 | 分子量: 107531.047 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: PEP3, VAM8, VPS18, VPT18, YLR148W, L9634.2 / 発現宿主: |
| #5: タンパク質 | 分子量: 123049.414 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: VAM6, CVT4, VPL18, VPL22, VPS39, YDL077C / 発現宿主: |
-Vacuolar protein sorting-associated protein ... , 3種, 3分子 BDF
| #2: タンパク質 | 分子量: 92857.000 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: VPS16, VAM9, VPT16, YPL045W / 発現宿主: |
|---|---|
| #4: タンパク質 | 分子量: 79354.977 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: VPS33, SLP1, VAM5, YLR396C, L8084.15 / 発現宿主: |
| #6: タンパク質 | 分子量: 116530.555 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: VPS41, FET2, VAM2, YDR080W, D446, YD8554.13 / 発現宿主: |
-詳細
| Has protein modification | N |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Tethering complex HOPS / タイプ: COMPLEX / Entity ID: all / 由来: RECOMBINANT |
|---|---|
| 由来(天然) | 生物種: |
| 由来(組換発現) | 生物種: |
| 緩衝液 | pH: 7.4 |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 顕微鏡 | モデル: TFS GLACIOS |
|---|---|
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 200 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 200 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 最大 デフォーカス(公称値): 2800 nm / 最小 デフォーカス(公称値): 800 nm |
| 撮影 | 電子線照射量: 50 e/Å2 フィルム・検出器のモデル: FEI FALCON IV (4k x 4k) |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: NONE | ||||||||||||||||||||||||
| 3次元再構成 | 解像度: 4.4 Å / 解像度の算出法: OTHER / 粒子像の数: 244661 詳細: Resolution of the consensus map of the lower part of the complex is used here 対称性のタイプ: POINT |
