ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
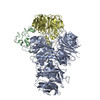
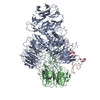
| 登録情報 | データベース: PDB / ID: 7zgq | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Polymerase module of yeast CPF in complex with the yPIM of Cft2 | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | RNA BINDING PROTEIN / CPF / 3'-end processing | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報: / Processing of Intronless Pre-mRNAs / termination of RNA polymerase II transcription, poly(A)-coupled / mRNA cleavage and polyadenylation specificity factor complex / mRNA 3'-end processing / termination of RNA polymerase II transcription / mRNA processing / mitochondrion / RNA binding / nucleus / metal ion binding 類似検索 - 分子機能 | |||||||||
| 生物種 |  | |||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 2.8 Å | |||||||||
 データ登録者 データ登録者 | Rodriguez-Molina, J.B. / Passmore, L.A. | |||||||||
| 資金援助 | European Union,  英国, 2件 英国, 2件
| |||||||||
 引用 引用 | ジャーナル: Mol Cell / 年: 2022 タイトル: Mpe1 senses the binding of pre-mRNA and controls 3' end processing by CPF. 著者: Juan B Rodríguez-Molina / Francis J O'Reilly / Holly Fagarasan / Eleanor Sheekey / Sarah Maslen / J Mark Skehel / Juri Rappsilber / Lori A Passmore /  要旨: Most eukaryotic messenger RNAs (mRNAs) are processed at their 3' end by the cleavage and polyadenylation specificity factor (CPF/CPSF). CPF mediates the endonucleolytic cleavage of the pre-mRNA and ...Most eukaryotic messenger RNAs (mRNAs) are processed at their 3' end by the cleavage and polyadenylation specificity factor (CPF/CPSF). CPF mediates the endonucleolytic cleavage of the pre-mRNA and addition of a polyadenosine (poly(A)) tail, which together define the 3' end of the mature transcript. The activation of CPF is highly regulated to maintain the fidelity of RNA processing. Here, using cryo-EM of yeast CPF, we show that the Mpe1 subunit directly contacts the polyadenylation signal sequence in nascent pre-mRNA. The region of Mpe1 that contacts RNA also promotes the activation of CPF endonuclease activity and controls polyadenylation. The Cft2 subunit of CPF antagonizes the RNA-stabilized configuration of Mpe1. In vivo, the depletion or mutation of Mpe1 leads to widespread defects in transcription termination by RNA polymerase II, resulting in transcription interference on neighboring genes. Together, our data suggest that Mpe1 plays a major role in accurate 3' end processing, activating CPF, and ensuring timely transcription termination. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7zgq.cif.gz | 372.3 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7zgq.ent.gz | 281.4 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 7zgq.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/zg/7zgqftp://data.pdbj.org/pub/pdb/validation_reports/zg/7zgq | HTTPS FTP |
|---|
-関連構造データ





-リンク
 PDBj
PDBj

- 集合体
集合体
| 登録構造単位 | 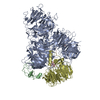
|
|---|---|
| 1 |
|
-要素
| #1: タンパク質 | 分子量: 80993.055 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: CFT2, YDH1, YLR115W, L2946 発現宿主:   Spodoptera frugiperda (ツマジロクサヨトウ) Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q12102 |
|---|---|
| #2: タンパク質 | 分子量: 153577.156 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 株: ATCC 204508 / S288c / 遺伝子: CFT1, YHH1, YDR301W 発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q06632 |
| #3: タンパク質 | 分子量: 24594.498 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: YTH1, GI527_G0006153 発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: A0A6A5Q2R8 |
| #4: タンパク質 | 分子量: 53211.117 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: PFS2, GI527_G0004795 発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: A0A6A5Q543 |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: polymerase module-Mpe1-Cft2(S)-RNA / タイプ: COMPLEX / Entity ID: all / 由来: RECOMBINANT |
|---|---|
| 分子量 | 実験値: NO |
| 由来(天然) | 生物種: |
| 由来(組換発現) | 生物種: Spodoptera frugiperda (ツマジロクサヨトウ) |
| 緩衝液 | pH: 8 / 詳細: 20 mM HEPES pH 8, 50 mM NaCl, 0.5 mM TCEP |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 試料支持 | グリッドの材料: GOLD / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: UltrAuFoil R1.2/1.3 |
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277.15 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 105000 X / 最大 デフォーカス(公称値): 3100 nm / 最小 デフォーカス(公称値): 500 nm / Cs: 2.7 mm / C2レンズ絞り径: 50 µm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN |
| 撮影 | 電子線照射量: 37 e/Å2 フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) 撮影したグリッド数: 1 / 実像数: 5118 |
| 電子光学装置 | エネルギーフィルタースリット幅: 20 eV |
- 解析
解析
| ソフトウェア | 名称: PHENIX / バージョン: 1.20_4459: / 分類: 精密化 | ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 1946027 | ||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 2.8 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 141584 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: OTHER / 空間: REAL | ||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 |
|