 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7nac | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | State E2 nucleolar 60S ribosomal biogenesis intermediate - Composite model | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | RIBOSOME / ribosome biogenesis / DEAD-box ATPases / methyltransferase / nucleolus | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報25S rRNA (cytosine2870-C5)-methyltransferase / 27S pre-rRNA (guanosine2922-2'-O)-methyltransferase / rRNA (guanosine-2'-O-)-methyltransferase activity / Noc1p-Noc2p complex / snoRNA release from pre-rRNA / Noc2p-Noc3p complex / nuclear division / exonucleolytic trimming to generate mature 5'-end of 5.8S rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / endonucleolytic cleavage in ITS1 upstream of 5.8S rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / rRNA (uridine-2'-O-)-methyltransferase activity ...25S rRNA (cytosine2870-C5)-methyltransferase / 27S pre-rRNA (guanosine2922-2'-O)-methyltransferase / rRNA (guanosine-2'-O-)-methyltransferase activity / Noc1p-Noc2p complex / snoRNA release from pre-rRNA / Noc2p-Noc3p complex / nuclear division / exonucleolytic trimming to generate mature 5'-end of 5.8S rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / endonucleolytic cleavage in ITS1 upstream of 5.8S rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / rRNA (uridine-2'-O-)-methyltransferase activity / rRNA (guanine) methyltransferase activity / rRNA (cytosine-C5-)-methyltransferase activity / endonucleolytic cleavage in 5'-ETS of tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / intracellular mRNA localization / endonucleolytic cleavage to generate mature 5'-end of SSU-rRNA from (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / pre-replicative complex assembly involved in nuclear cell cycle DNA replication / PeBoW complex / nuclear pre-replicative complex / rRNA base methylation / rRNA primary transcript binding / ATP-dependent activity, acting on RNA / positive regulation of ATP-dependent activity / maturation of 5.8S rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / rRNA methylation / protein folding chaperone complex / pre-mRNA 5'-splice site binding / cleavage in ITS2 between 5.8S rRNA and LSU-rRNA of tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / proteasome binding / maturation of 5.8S rRNA / Major pathway of rRNA processing in the nucleolus and cytosol / SRP-dependent cotranslational protein targeting to membrane / GTP hydrolysis and joining of the 60S ribosomal subunit / ribosomal large subunit binding / Nonsense Mediated Decay (NMD) independent of the Exon Junction Complex (EJC) / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / Formation of a pool of free 40S subunits / negative regulation of mRNA splicing, via spliceosome / ATPase activator activity / nuclear-transcribed mRNA catabolic process / preribosome, large subunit precursor / L13a-mediated translational silencing of Ceruloplasmin expression / translational elongation / ribosomal large subunit export from nucleus / mRNA transport / DNA replication initiation / 90S preribosome / regulation of translational fidelity / protein-RNA complex assembly / ribosomal subunit export from nucleus / ribonucleoprotein complex binding / endonucleolytic cleavage in ITS1 to separate SSU-rRNA from 5.8S rRNA and LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / translation initiation factor activity / maturation of LSU-rRNA / proteasome complex / maturation of LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / nuclear periphery / assembly of large subunit precursor of preribosome / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / ribosomal large subunit biogenesis / cytosolic ribosome assembly / maturation of SSU-rRNA / small-subunit processome / macroautophagy / maintenance of translational fidelity / protein catabolic process / rRNA processing / transcription corepressor activity / ribosomal small subunit biogenesis / protein transport / ribosome biogenesis / ribosomal large subunit assembly / nuclear envelope / protein-macromolecule adaptor activity / histone binding / ATPase binding / large ribosomal subunit rRNA binding / 5S rRNA binding / cytosolic large ribosomal subunit / cytoplasmic translation / RNA helicase activity / negative regulation of translation / rRNA binding / ribosome / RNA helicase / structural constituent of ribosome / translation / cell division / GTPase activity / mRNA binding / chromatin binding / GTP binding / nucleolus / negative regulation of transcription by RNA polymerase II / ATP hydrolysis activity / DNA binding / RNA binding / nucleoplasm / ATP binding / identical protein binding / nucleus 類似検索 - 分子機能 | ||||||||||||
| 生物種 |  | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.04 Å | ||||||||||||
 データ登録者 データ登録者 | Cruz, V.E. / Sekulski, K. / Peddada, N. / Erzberger, J.P. | ||||||||||||
| 資金援助 |  米国, 3件 米国, 3件
| ||||||||||||
 引用 引用 | ジャーナル: Nat Struct Mol Biol / 年: 2022 タイトル: Sequence-specific remodeling of a topologically complex RNP substrate by Spb4. 著者: Victor Emmanuel Cruz / Kamil Sekulski / Nagesh Peddada / Carolin Sailer / Sahana Balasubramanian / Christine S Weirich / Florian Stengel / Jan P Erzberger /   要旨: DEAD-box ATPases are ubiquitous enzymes essential in all aspects of RNA biology. However, the limited in vitro catalytic activities described for these enzymes are at odds with their complex cellular ...DEAD-box ATPases are ubiquitous enzymes essential in all aspects of RNA biology. However, the limited in vitro catalytic activities described for these enzymes are at odds with their complex cellular roles, most notably in driving large-scale RNA remodeling steps during the assembly of ribonucleoproteins (RNPs). We describe cryo-EM structures of 60S ribosomal biogenesis intermediates that reveal how context-specific RNA unwinding by the DEAD-box ATPase Spb4 results in extensive, sequence-specific remodeling of rRNA secondary structure. Multiple cis and trans interactions stabilize Spb4 in a post-catalytic, high-energy intermediate that drives the organization of the three-way junction at the base of rRNA domain IV. This mechanism explains how limited strand separation by DEAD-box ATPases is leveraged to provide non-equilibrium directionality and ensure efficient and accurate RNP assembly. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7nac.cif.gz | 3.5 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7nac.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 7nac.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 7nac_validation.pdf.gz | 2 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 7nac_full_validation.pdf.gz | 2.1 MB | 表示 | |
| XML形式データ | 7nac_validation.xml.gz | 307.8 KB | 表示 | |
| CIF形式データ | 7nac_validation.cif.gz | 524.1 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/na/7nacftp://data.pdbj.org/pub/pdb/validation_reports/na/7nac | HTTPS FTP |
-関連構造データ
| 関連構造データ |  24269MC  7nadC  7nafC  7r6kC  7r6qC  7r72C  7r7aC 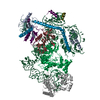 7r7cC  7u0hC M: このデータのモデリングに利用したマップデータ C: 同じ文献を引用 ( |
|---|---|
| 類似構造データ |







-リンク
 PDBj
PDBj

































- 集合体
集合体
| 登録構造単位 | 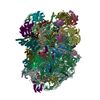
|
|---|---|
| 1 |
|
-要素
-RNA鎖 , 3種, 3分子 126
| #1: RNA鎖 | 分子量: 1097574.875 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 |
|---|---|
| #2: RNA鎖 | 分子量: 50682.922 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 |
| #52: RNA鎖 | 分子量: 27786.307 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 |
-タンパク質 , 18種, 18分子 7JWblqsvyz8IKnpwa5
| #3: タンパク質 | 分子量: 23652.645 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: P43586 |
|---|---|
| #10: タンパク質 | 分子量: 49842.613 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: P36049 |
| #19: タンパク質 | 分子量: 27098.012 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: P33201 |
| #22: タンパク質 | 分子量: 74531.227 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: Q02892 |
| #29: タンパク質 | 分子量: 20411.699 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: Q08962 |
| #31: タンパク質 | 分子量: 69912.164 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 参照: UniProt: P40991, 25S rRNA (cytosine2870-C5)-methyltransferase |
| #33: タンパク質 | 分子量: 57798.652 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: P40010 |
| #35: タンパク質 | 分子量: 26954.447 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: P40007 |
| #36: タンパク質 | 分子量: 26476.605 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: Q12522 |
| #37: タンパク質 | 分子量: 12435.429 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: P38202 |
| #38: タンパク質 | 分子量: 81719.289 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: A0A8H4BX61 |
| #39: タンパク質 | 分子量: 75689.008 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: Q07896 |
| #40: タンパク質 | 分子量: 42596.691 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: P38779 |
| #49: タンパク質 | 分子量: 69984.148 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: P53261 |
| #50: タンパク質 | 分子量: 51426.637 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: A0A8H4BV53 |
| #55: タンパク質 | 分子量: 96656.172 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 参照: UniProt: P25582, 27S pre-rRNA (guanosine2922-2'-O)-methyltransferase |
| #56: タンパク質 | 分子量: 24524.799 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: A0A6A5Q2N7 |
| #57: タンパク質 | 分子量: 28032.553 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: A0A6A5Q2X9 |
-Ribosome biogenesis protein ... , 6種, 6分子 Amruto
| #4: タンパク質 | 分子量: 33585.414 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: Q08235 |
|---|---|
| #30: タンパク質 | 分子量: 91772.570 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: Q04660 |
| #32: タンパク質 | 分子量: 29786.783 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: A6ZR80 |
| #34: タンパク質 | 分子量: 24027.650 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: Q07915 |
| #51: タンパク質 | 分子量: 36621.074 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: P40693 |
| #54: タンパク質 | 分子量: 25499.186 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: P53927 |
+60S ribosomal protein ... , 29種, 29分子 BCEFHLMNOPQSTXYdefhijVRGZUcgk
-ATP-dependent ... , 2種, 2分子 Dx
| #53: タンパク質 | 分子量: 56798.348 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: Q03532, RNA helicase |
|---|---|
| #58: タンパク質 | 分子量: 69492.172 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: BY4741 / 参照: UniProt: A7A237, RNA helicase |
-非ポリマー , 2種, 3分子 
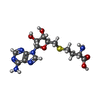
| #59: 化合物 |  分子量: 65.409 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Zn#60: 化合物 | ChemComp-SAH / |  タイプ: L-peptide linking / 分子量: 384.411 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C14H20N6O5S / タイプ: SUBJECT OF INVESTIGATION タイプ: L-peptide linking / 分子量: 384.411 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C14H20N6O5S / タイプ: SUBJECT OF INVESTIGATION |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Nucleolar 60S intermediate purified with tags on Ytm1 and Spb4. タイプ: RIBOSOME / Entity ID: #1-#58 / 由来: NATURAL | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: 3.3 MDa / 実験値: NO | ||||||||||||||||||||||||||||||
| 由来(天然) | 生物種: | ||||||||||||||||||||||||||||||
| 緩衝液 | pH: 8 | ||||||||||||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||||||||||||
| 試料 | 濃度: 0.45 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||
| 試料支持 | グリッドの材料: COPPER / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: Quantifoil R2/2 | ||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 81000 X / 最大 デフォーカス(公称値): 2200 nm / 最小 デフォーカス(公称値): 900 nm / C2レンズ絞り径: 70 µm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 平均露光時間: 0.05 sec. / 電子線照射量: 1.2 e/Å2 / フィルム・検出器のモデル: GATAN K3 (6k x 4k) / 撮影したグリッド数: 1 / 実像数: 4523 |
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||
| CTF補正 | タイプ: NONE | ||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 825096 | ||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.04 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 198000 / アルゴリズム: FOURIER SPACE / クラス平均像の数: 1 / 対称性のタイプ: POINT | ||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT | ||||||||||||||||||||||||
| 精密化 | 交差検証法: NONE 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 14.55 Å2 | ||||||||||||||||||||||||
| 拘束条件 |
|
