 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 5l4k | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | The human 26S proteasome lid | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | STRUCTURAL PROTEIN / proteostasis / AAA-ATPase | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報Impaired BRCA2 translocation to the nucleus / Impaired BRCA2 binding to SEM1 (DSS1) / meiosis I / 加水分解酵素; プロテアーゼ; ペプチド結合加水分解酵素; オメガペプチターゼ / proteasome accessory complex / integrator complex / proteasome regulatory particle / cytosolic proteasome complex / proteasome regulatory particle, lid subcomplex / proteasome regulatory particle, base subcomplex ...Impaired BRCA2 translocation to the nucleus / Impaired BRCA2 binding to SEM1 (DSS1) / meiosis I / 加水分解酵素; プロテアーゼ; ペプチド結合加水分解酵素; オメガペプチターゼ / proteasome accessory complex / integrator complex / proteasome regulatory particle / cytosolic proteasome complex / proteasome regulatory particle, lid subcomplex / proteasome regulatory particle, base subcomplex / metal-dependent deubiquitinase activity / protein K63-linked deubiquitination / Homologous DNA Pairing and Strand Exchange / Defective homologous recombination repair (HRR) due to BRCA1 loss of function / Defective HDR through Homologous Recombination Repair (HRR) due to PALB2 loss of BRCA1 binding function / Defective HDR through Homologous Recombination Repair (HRR) due to PALB2 loss of BRCA2/RAD51/RAD51C binding function / Resolution of D-loop Structures through Synthesis-Dependent Strand Annealing (SDSA) / Regulation of ornithine decarboxylase (ODC) / Resolution of D-loop Structures through Holliday Junction Intermediates / Cross-presentation of soluble exogenous antigens (endosomes) / K63-linked deubiquitinase activity / Somitogenesis / Impaired BRCA2 binding to RAD51 / proteasome binding / regulation of protein catabolic process / proteasome storage granule / Presynaptic phase of homologous DNA pairing and strand exchange / endopeptidase activator activity / polyubiquitin modification-dependent protein binding / proteasome assembly / protein deubiquitination / mRNA export from nucleus / regulation of proteasomal protein catabolic process / enzyme regulator activity / proteasome complex / Regulation of activated PAK-2p34 by proteasome mediated degradation / Autodegradation of Cdh1 by Cdh1:APC/C / APC/C:Cdc20 mediated degradation of Securin / Asymmetric localization of PCP proteins / SCF-beta-TrCP mediated degradation of Emi1 / NIK-->noncanonical NF-kB signaling / Ubiquitin-dependent degradation of Cyclin D / AUF1 (hnRNP D0) binds and destabilizes mRNA / TNFR2 non-canonical NF-kB pathway / stem cell differentiation / Vpu mediated degradation of CD4 / Assembly of the pre-replicative complex / Degradation of DVL / Ubiquitin Mediated Degradation of Phosphorylated Cdc25A / Cdc20:Phospho-APC/C mediated degradation of Cyclin A / Dectin-1 mediated noncanonical NF-kB signaling / Hh mutants are degraded by ERAD / Degradation of AXIN / Activation of NF-kappaB in B cells / Degradation of GLI1 by the proteasome / Hedgehog ligand biogenesis / Defective CFTR causes cystic fibrosis / G2/M Checkpoints / Negative regulation of NOTCH4 signaling / GSK3B and BTRC:CUL1-mediated-degradation of NFE2L2 / Autodegradation of the E3 ubiquitin ligase COP1 / Vif-mediated degradation of APOBEC3G / double-strand break repair via homologous recombination / Regulation of RUNX3 expression and activity / Hedgehog 'on' state / Degradation of GLI2 by the proteasome / GLI3 is processed to GLI3R by the proteasome / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / MAPK6/MAPK4 signaling / APC/C:Cdh1 mediated degradation of Cdc20 and other APC/C:Cdh1 targeted proteins in late mitosis/early G1 / Degradation of beta-catenin by the destruction complex / ABC-family proteins mediated transport / Oxygen-dependent proline hydroxylation of Hypoxia-inducible Factor Alpha / HDR through Homologous Recombination (HRR) / CDK-mediated phosphorylation and removal of Cdc6 / Metalloprotease DUBs / CLEC7A (Dectin-1) signaling / SCF(Skp2)-mediated degradation of p27/p21 / double-strand break repair via nonhomologous end joining / Regulation of expression of SLITs and ROBOs / FCERI mediated NF-kB activation / Regulation of PTEN stability and activity / Interleukin-1 signaling / Orc1 removal from chromatin / Regulation of RAS by GAPs / Separation of Sister Chromatids / Regulation of RUNX2 expression and activity / The role of GTSE1 in G2/M progression after G2 checkpoint / metallopeptidase activity / azurophil granule lumen / UCH proteinases / KEAP1-NFE2L2 pathway / Antigen processing: Ubiquitination & Proteasome degradation / Downstream TCR signaling / RUNX1 regulates transcription of genes involved in differentiation of HSCs / Neddylation / ER-Phagosome pathway / ubiquitin-dependent protein catabolic process / secretory granule lumen / proteasome-mediated ubiquitin-dependent protein catabolic process 類似検索 - 分子機能 | ||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.9 Å | ||||||
 データ登録者 データ登録者 | Schweitzer, A. / Aufderheide, A. / Rudack, T. / Beck, F. | ||||||
 引用 引用 | ジャーナル: Proc Natl Acad Sci U S A / 年: 2016 タイトル: Structure of the human 26S proteasome at a resolution of 3.9 Å. 著者: Andreas Schweitzer / Antje Aufderheide / Till Rudack / Florian Beck / Günter Pfeifer / Jürgen M Plitzko / Eri Sakata / Klaus Schulten / Friedrich Förster / Wolfgang Baumeister /    要旨: Protein degradation in eukaryotic cells is performed by the Ubiquitin-Proteasome System (UPS). The 26S proteasome holocomplex consists of a core particle (CP) that proteolytically degrades ...Protein degradation in eukaryotic cells is performed by the Ubiquitin-Proteasome System (UPS). The 26S proteasome holocomplex consists of a core particle (CP) that proteolytically degrades polyubiquitylated proteins, and a regulatory particle (RP) containing the AAA-ATPase module. This module controls access to the proteolytic chamber inside the CP and is surrounded by non-ATPase subunits (Rpns) that recognize substrates and deubiquitylate them before unfolding and degradation. The architecture of the 26S holocomplex is highly conserved between yeast and humans. The structure of the human 26S holocomplex described here reveals previously unidentified features of the AAA-ATPase heterohexamer. One subunit, Rpt6, has ADP bound, whereas the other five have ATP in their binding pockets. Rpt6 is structurally distinct from the other five Rpt subunits, most notably in its pore loop region. For Rpns, the map reveals two main, previously undetected, features: the C terminus of Rpn3 protrudes into the mouth of the ATPase ring; and Rpn1 and Rpn2, the largest proteasome subunits, are linked by an extended connection. The structural features of the 26S proteasome observed in this study are likely to be important for coordinating the proteasomal subunits during substrate processing. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 5l4k.cif.gz | 1.8 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb5l4k.ent.gz | 1.5 MB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 5l4k.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 5l4k_validation.pdf.gz | 858.2 KB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 5l4k_full_validation.pdf.gz | 908.1 KB | 表示 | |
| XML形式データ | 5l4k_validation.xml.gz | 122.1 KB | 表示 | |
| CIF形式データ | 5l4k_validation.cif.gz | 190.8 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/l4/5l4kftp://data.pdbj.org/pub/pdb/validation_reports/l4/5l4k | HTTPS FTP |
-関連構造データ


-リンク
 PDBj
PDBj







- 集合体
集合体
| 登録構造単位 | 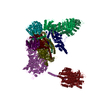
|
|---|---|
| 1 |
|
-要素
-26S proteasome non-ATPase regulatory subunit ... , 11種, 11分子 WVTZNSPQRUO
| #1: タンパク質 | 分子量: 40781.590 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: P55036 |
|---|---|
| #2: タンパク質 | 分子量: 34620.023 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト)参照: UniProt: O00487, 加水分解酵素; プロテアーゼ; ペプチド結合加水分解酵素; オメガペプチターゼ |
| #3: タンパク質 | 分子量: 39667.871 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: P48556 |
| #5: タンパク質 | 分子量: 100313.625 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: Q13200 |
| #6: タンパク質 | 分子量: 105958.234 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: Q99460 |
| #7: タンパク質 | 分子量: 61066.500 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: O43242 |
| #8: タンパク質 | 分子量: 52979.359 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: O00232 |
| #9: タンパク質 | 分子量: 47526.688 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: O00231 |
| #10: タンパク質 | 分子量: 45592.285 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: Q15008 |
| #11: タンパク質 | 分子量: 37086.441 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: P51665 |
| #12: タンパク質 | 分子量: 42995.359 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: Q9UNM6 |
-タンパク質 , 1種, 1分子 Y
| #4: タンパク質 | 分子量: 8284.611 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Homo sapiens (ヒト) / 参照: UniProt: P60896 |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: human 26S proteasome lid / タイプ: COMPLEX / Entity ID: all / 由来: NATURAL |
|---|---|
| 分子量 | 値: 2.5 MDa / 実験値: NO |
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 緩衝液 | pH: 7.5 |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES / 詳細: This sample was monodisperse. |
| 試料支持 | グリッドの材料: COPPER / グリッドのサイズ: 200 divisions/in. / グリッドのタイプ: Quantifoil R2/1 |
| 急速凍結 | 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD |
| 撮影 | 平均露光時間: 1.5 sec. / 電子線照射量: 45 e/Å2 / フィルム・検出器のモデル: OTHER / 撮影したグリッド数: 19 / 実像数: 40211 |
| 画像スキャン | 横: 4096 / 縦: 4096 |
- 解析
解析
| ソフトウェア | 名称: PHENIX / バージョン: (1.10.1_2155: ???) / 分類: 精密化 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 画像処理 | 詳細: Falcon III | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 688742 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.9 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 461402 / アルゴリズム: FOURIER SPACE / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: FLEXIBLE FIT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 | 解像度: 3.9→3.9 Å / SU ML: 1.03 / σ(F): 1.57 / 位相誤差: 46.21 / 立体化学のターゲット値: MLHL
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | 減衰半径: 0.9 Å / VDWプローブ半径: 1.11 Å / 溶媒モデル: FLAT BULK SOLVENT MODEL | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLS | 手法: refined / Refine-ID: ELECTRON MICROSCOPY
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 TLSグループ |
|