 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 5l4k | ||||||
|---|---|---|---|---|---|---|---|
| Title | The human 26S proteasome lid | ||||||
 Components Components |
| ||||||
 Keywords Keywords | STRUCTURAL PROTEIN / proteostasis / AAA-ATPase | ||||||
| Function / homology |  Function and homology information Function and homology informationImpaired BRCA2 translocation to the nucleus / Impaired BRCA2 binding to SEM1 (DSS1) / meiosis I / Hydrolases; Acting on peptide bonds (peptidases); Omega peptidases / proteasome accessory complex / integrator complex / proteasome regulatory particle / cytosolic proteasome complex / proteasome regulatory particle, lid subcomplex / proteasome regulatory particle, base subcomplex ...Impaired BRCA2 translocation to the nucleus / Impaired BRCA2 binding to SEM1 (DSS1) / meiosis I / Hydrolases; Acting on peptide bonds (peptidases); Omega peptidases / proteasome accessory complex / integrator complex / proteasome regulatory particle / cytosolic proteasome complex / proteasome regulatory particle, lid subcomplex / proteasome regulatory particle, base subcomplex / metal-dependent deubiquitinase activity / protein K63-linked deubiquitination / Homologous DNA Pairing and Strand Exchange / Defective homologous recombination repair (HRR) due to BRCA1 loss of function / Defective HDR through Homologous Recombination Repair (HRR) due to PALB2 loss of BRCA1 binding function / Defective HDR through Homologous Recombination Repair (HRR) due to PALB2 loss of BRCA2/RAD51/RAD51C binding function / Resolution of D-loop Structures through Synthesis-Dependent Strand Annealing (SDSA) / Regulation of ornithine decarboxylase (ODC) / Resolution of D-loop Structures through Holliday Junction Intermediates / Cross-presentation of soluble exogenous antigens (endosomes) / K63-linked deubiquitinase activity / Somitogenesis / Impaired BRCA2 binding to RAD51 / proteasome binding / regulation of protein catabolic process / proteasome storage granule / Presynaptic phase of homologous DNA pairing and strand exchange / endopeptidase activator activity / polyubiquitin modification-dependent protein binding / proteasome assembly / protein deubiquitination / mRNA export from nucleus / regulation of proteasomal protein catabolic process / enzyme regulator activity / proteasome complex / Regulation of activated PAK-2p34 by proteasome mediated degradation / Autodegradation of Cdh1 by Cdh1:APC/C / APC/C:Cdc20 mediated degradation of Securin / Asymmetric localization of PCP proteins / SCF-beta-TrCP mediated degradation of Emi1 / NIK-->noncanonical NF-kB signaling / Ubiquitin-dependent degradation of Cyclin D / AUF1 (hnRNP D0) binds and destabilizes mRNA / TNFR2 non-canonical NF-kB pathway / stem cell differentiation / Vpu mediated degradation of CD4 / Assembly of the pre-replicative complex / Degradation of DVL / Ubiquitin Mediated Degradation of Phosphorylated Cdc25A / Cdc20:Phospho-APC/C mediated degradation of Cyclin A / Dectin-1 mediated noncanonical NF-kB signaling / Hh mutants are degraded by ERAD / Degradation of AXIN / Activation of NF-kappaB in B cells / Degradation of GLI1 by the proteasome / Hedgehog ligand biogenesis / Defective CFTR causes cystic fibrosis / G2/M Checkpoints / Negative regulation of NOTCH4 signaling / GSK3B and BTRC:CUL1-mediated-degradation of NFE2L2 / Autodegradation of the E3 ubiquitin ligase COP1 / Vif-mediated degradation of APOBEC3G / double-strand break repair via homologous recombination / Regulation of RUNX3 expression and activity / Hedgehog 'on' state / Degradation of GLI2 by the proteasome / GLI3 is processed to GLI3R by the proteasome / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / MAPK6/MAPK4 signaling / APC/C:Cdh1 mediated degradation of Cdc20 and other APC/C:Cdh1 targeted proteins in late mitosis/early G1 / Degradation of beta-catenin by the destruction complex / ABC-family proteins mediated transport / Oxygen-dependent proline hydroxylation of Hypoxia-inducible Factor Alpha / HDR through Homologous Recombination (HRR) / CDK-mediated phosphorylation and removal of Cdc6 / Metalloprotease DUBs / CLEC7A (Dectin-1) signaling / SCF(Skp2)-mediated degradation of p27/p21 / double-strand break repair via nonhomologous end joining / Regulation of expression of SLITs and ROBOs / FCERI mediated NF-kB activation / Regulation of PTEN stability and activity / Interleukin-1 signaling / Orc1 removal from chromatin / Regulation of RAS by GAPs / Separation of Sister Chromatids / Regulation of RUNX2 expression and activity / The role of GTSE1 in G2/M progression after G2 checkpoint / metallopeptidase activity / azurophil granule lumen / UCH proteinases / KEAP1-NFE2L2 pathway / Antigen processing: Ubiquitination & Proteasome degradation / Downstream TCR signaling / RUNX1 regulates transcription of genes involved in differentiation of HSCs / Neddylation / ER-Phagosome pathway / ubiquitin-dependent protein catabolic process / secretory granule lumen / proteasome-mediated ubiquitin-dependent protein catabolic process Similarity search - Function | ||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.9 Å | ||||||
 Authors Authors | Schweitzer, A. / Aufderheide, A. / Rudack, T. / Beck, F. | ||||||
 Citation Citation | Journal: Proc Natl Acad Sci U S A / Year: 2016 Title: Structure of the human 26S proteasome at a resolution of 3.9 Å. Authors: Andreas Schweitzer / Antje Aufderheide / Till Rudack / Florian Beck / Günter Pfeifer / Jürgen M Plitzko / Eri Sakata / Klaus Schulten / Friedrich Förster / Wolfgang Baumeister /    Abstract: Protein degradation in eukaryotic cells is performed by the Ubiquitin-Proteasome System (UPS). The 26S proteasome holocomplex consists of a core particle (CP) that proteolytically degrades ...Protein degradation in eukaryotic cells is performed by the Ubiquitin-Proteasome System (UPS). The 26S proteasome holocomplex consists of a core particle (CP) that proteolytically degrades polyubiquitylated proteins, and a regulatory particle (RP) containing the AAA-ATPase module. This module controls access to the proteolytic chamber inside the CP and is surrounded by non-ATPase subunits (Rpns) that recognize substrates and deubiquitylate them before unfolding and degradation. The architecture of the 26S holocomplex is highly conserved between yeast and humans. The structure of the human 26S holocomplex described here reveals previously unidentified features of the AAA-ATPase heterohexamer. One subunit, Rpt6, has ADP bound, whereas the other five have ATP in their binding pockets. Rpt6 is structurally distinct from the other five Rpt subunits, most notably in its pore loop region. For Rpns, the map reveals two main, previously undetected, features: the C terminus of Rpn3 protrudes into the mouth of the ATPase ring; and Rpn1 and Rpn2, the largest proteasome subunits, are linked by an extended connection. The structural features of the 26S proteasome observed in this study are likely to be important for coordinating the proteasomal subunits during substrate processing. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 5l4k.cif.gz | 1.8 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb5l4k.ent.gz | 1.5 MB | Display | PDB format |
| PDBx/mmJSON format | 5l4k.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Summary document | 5l4k_validation.pdf.gz | 858.2 KB | Display | wwPDB validaton report |
|---|---|---|---|---|
| Full document | 5l4k_full_validation.pdf.gz | 908.1 KB | Display | |
| Data in XML | 5l4k_validation.xml.gz | 122.1 KB | Display | |
| Data in CIF | 5l4k_validation.cif.gz | 190.8 KB | Display | |
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/l4/5l4kftp://data.pdbj.org/pub/pdb/validation_reports/l4/5l4k | HTTPS FTP |
-Related structure data
| Related structure data |  4002MC  5l4gC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |


-Links
 PDBj
PDBj







- Assembly
Assembly
| Deposited unit | 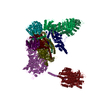
|
|---|---|
| 1 |
|
-Components
-26S proteasome non-ATPase regulatory subunit ... , 11 types, 11 molecules WVTZNSPQRUO
| #1: Protein | Mass: 40781.590 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P55036 |
|---|---|
| #2: Protein | Mass: 34620.023 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human)References: UniProt: O00487, Hydrolases; Acting on peptide bonds (peptidases); Omega peptidases |
| #3: Protein | Mass: 39667.871 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P48556 |
| #5: Protein | Mass: 100313.625 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: Q13200 |
| #6: Protein | Mass: 105958.234 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: Q99460 |
| #7: Protein | Mass: 61066.500 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: O43242 |
| #8: Protein | Mass: 52979.359 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: O00232 |
| #9: Protein | Mass: 47526.688 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: O00231 |
| #10: Protein | Mass: 45592.285 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: Q15008 |
| #11: Protein | Mass: 37086.441 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P51665 |
| #12: Protein | Mass: 42995.359 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: Q9UNM6 |
-Protein , 1 types, 1 molecules Y
| #4: Protein | Mass: 8284.611 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P60896 |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: human 26S proteasome lid / Type: COMPLEX / Entity ID: all / Source: NATURAL |
|---|---|
| Molecular weight | Value: 2.5 MDa / Experimental value: NO |
| Source (natural) | Organism: Homo sapiens (human) |
| Buffer solution | pH: 7.5 |
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES / Details: This sample was monodisperse. |
| Specimen support | Grid material: COPPER / Grid mesh size: 200 divisions/in. / Grid type: Quantifoil R2/1 |
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD |
| Image recording | Average exposure time: 1.5 sec. / Electron dose: 45 e/Å2 / Film or detector model: OTHER / Num. of grids imaged: 19 / Num. of real images: 40211 |
| Image scans | Width: 4096 / Height: 4096 |
- Processing
Processing
| Software | Name: PHENIX / Version: (1.10.1_2155: ???) / Classification: refinement | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Image processing | Details: Falcon III | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 688742 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.9 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 461402 / Algorithm: FOURIER SPACE / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building | Protocol: FLEXIBLE FIT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement | Resolution: 3.9→3.9 Å / SU ML: 1.03 / σ(F): 1.57 / Phase error: 46.21 / Stereochemistry target values: MLHL
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.11 Å / Solvent model: FLAT BULK SOLVENT MODEL | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS params. | Method: refined / Refine-ID: ELECTRON MICROSCOPY
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS group |
|