Nucleosome / Complex / NUCLEAR PROTEIN / NUCLEAR PROTEIN-DNA complex
機能・相同性
機能・相同性情報
protein localization to CENP-A containing chromatin / CENP-A containing nucleosome / negative regulation of tumor necrosis factor-mediated signaling pathway / Replacement of protamines by nucleosomes in the male pronucleus / arachidonate 15-lipoxygenase / arachidonate 15-lipoxygenase activity / Packaging Of Telomere Ends / lipoxygenase pathway / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine ...protein localization to CENP-A containing chromatin / CENP-A containing nucleosome / negative regulation of tumor necrosis factor-mediated signaling pathway / Replacement of protamines by nucleosomes in the male pronucleus / arachidonate 15-lipoxygenase / arachidonate 15-lipoxygenase activity / Packaging Of Telomere Ends / lipoxygenase pathway / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / telomere organization / arachidonate metabolic process / Chromatin modifying enzymes / lipid oxidation / Deposition of new CENPA-containing nucleosomes at the centromere / hepoxilin biosynthetic process / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / linoleic acid metabolic process / Meiotic synapsis / Inhibition of DNA recombination at telomere / nucleosomal DNA binding / RNA Polymerase I Promoter Opening / Assembly of the ORC complex at the origin of replication / Interleukin-7 signaling / epigenetic regulation of gene expression / DNA methylation / HCMV Late Events / Condensation of Prophase Chromosomes / SIRT1 negatively regulates rRNA expression / Chromatin modifications during the maternal to zygotic transition (MZT) / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / PRC2 methylates histones and DNA / Defective pyroptosis / Meiotic recombination / innate immune response in mucosa / DNA Damage/Telomere Stress Induced Senescence / HDACs deacetylate histones / Nonhomologous End-Joining (NHEJ) / RNA Polymerase I Promoter Escape / Transcriptional regulation by small RNAs / lipopolysaccharide binding / Transcriptional regulation of granulopoiesis / HDMs demethylate histones / Formation of the beta-catenin:TCF transactivating complex / HCMV Early Events / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / G2/M DNA damage checkpoint / NoRC negatively regulates rRNA expression / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / PKMTs methylate histone lysines / B-WICH complex positively regulates rRNA expression / heterochromatin formation / RMTs methylate histone arginines / Pre-NOTCH Transcription and Translation / Metalloprotease DUBs / Activation of anterior HOX genes in hindbrain development during early embryogenesis / structural constituent of chromatin / UCH proteinases / nucleosome / antimicrobial humoral immune response mediated by antimicrobial peptide / Factors involved in megakaryocyte development and platelet production / Processing of DNA double-strand break ends / nucleosome assembly / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / chromatin organization / E3 ubiquitin ligases ubiquitinate target proteins / Senescence-Associated Secretory Phenotype (SASP) / RUNX1 regulates transcription of genes involved in differentiation of HSCs / HATs acetylate histones / gene expression / antibacterial humoral response / Oxidative Stress Induced Senescence / defense response to Gram-negative bacterium / Estrogen-dependent gene expression / killing of cells of another organism / Ub-specific processing proteases / defense response to Gram-positive bacterium / cadherin binding / protein heterodimerization activity / Amyloid fiber formation / negative regulation of cell population proliferation / protein-containing complex / DNA binding / extracellular space / extracellular exosome / extracellular region / nucleoplasm / membrane / nucleus / metal ion binding / cytosol 類似検索 - 分子機能
Japan Agency for Medical Research and Development (AMED)
JP22ama121009
日本
Japan Science and Technology
JPMJER1901
日本
Japan Society for the Promotion of Science (JSPS)
JP20K06599
日本
Japan Society for the Promotion of Science (JSPS)
JP21H05154
日本
Japan Science and Technology
JPMJPR18K9
日本
引用
ジャーナル: Commun Biol / 年: 2024 タイトル: Genome-wide mapping and cryo-EM structural analyses of the overlapping tri-nucleosome composed of hexasome-hexasome-octasome moieties. 著者: Masahiro Nishimura / Takeru Fujii / Hiroki Tanaka / Kazumitsu Maehara / Ken Morishima / Masahiro Shimizu / Yuki Kobayashi / Kayo Nozawa / Yoshimasa Takizawa / Masaaki Sugiyama / Yasuyuki ...著者: Masahiro Nishimura / Takeru Fujii / Hiroki Tanaka / Kazumitsu Maehara / Ken Morishima / Masahiro Shimizu / Yuki Kobayashi / Kayo Nozawa / Yoshimasa Takizawa / Masaaki Sugiyama / Yasuyuki Ohkawa / Hitoshi Kurumizaka / 要旨: The nucleosome is a fundamental unit of chromatin in which about 150 base pairs of DNA are wrapped around a histone octamer. The overlapping di-nucleosome has been proposed as a product of chromatin ...The nucleosome is a fundamental unit of chromatin in which about 150 base pairs of DNA are wrapped around a histone octamer. The overlapping di-nucleosome has been proposed as a product of chromatin remodeling around the transcription start site, and previously found as a chromatin unit, in which about 250 base pairs of DNA continuously bind to the histone core composed of a hexamer and an octamer. In the present study, our genome-wide analysis of human cells suggests another higher nucleosome stacking structure, the overlapping tri-nucleosome, which wraps about 300-350 base-pairs of DNA in the region downstream of certain transcription start sites of actively transcribed genes. We determine the cryo-electron microscopy (cryo-EM) structure of the overlapping tri-nucleosome, in which three subnucleosome moieties, hexasome, hexasome, and octasome, are associated by short connecting DNA segments. Small angle X-ray scattering and coarse-grained molecular dynamics simulation analyses reveal that the cryo-EM structure of the overlapping tri-nucleosome may reflect its structure in solution. Our findings suggest that nucleosome stacking structures composed of hexasome and octasome moieties may be formed by nucleosome remodeling factors around transcription start sites for gene regulation.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報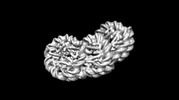
 マップデータ
マップデータ 試料
試料 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト) / synthetic construct (人工物)
Homo sapiens (ヒト) / synthetic construct (人工物) データ登録者
データ登録者 日本, 8件
日本, 8件  引用
引用
 構造の表示
構造の表示
 ダウンロードとリンク
ダウンロードとリンク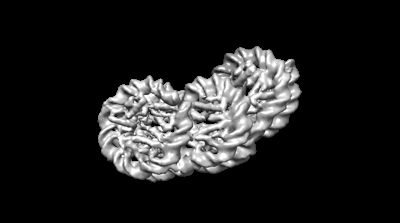 emd_35448.png
emd_35448.png http://ftp.pdbj.org/pub/emdb/structures/EMD-35448
http://ftp.pdbj.org/pub/emdb/structures/EMD-35448









 Z
Z Y
Y X
X








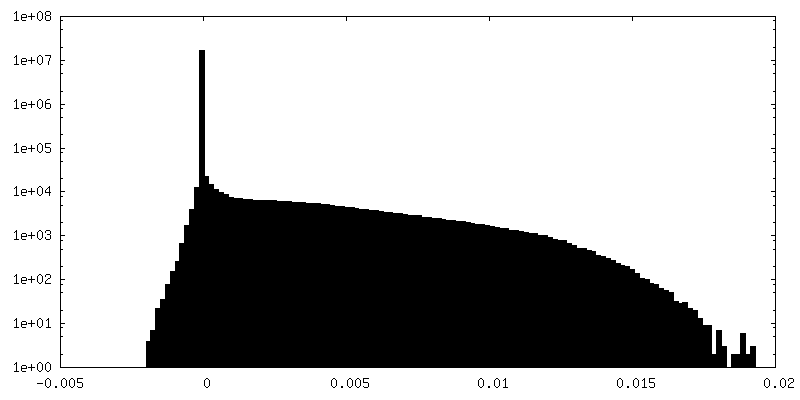






















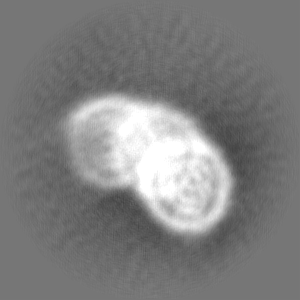












 試料の構成要素
試料の構成要素
 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN

