 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 5oa3 | ||||||
|---|---|---|---|---|---|---|---|
| Title | Human 40S-eIF2D-re-initiation complex | ||||||
 Components Components |
| ||||||
 Keywords Keywords | TRANSLATION / translation re-initiation complex / small ribosomal subunit / RNA binding protein / eukaryotic translation initiation factor | ||||||
| Function / homology |  Function and homology information Function and homology informationIRES-dependent viral translational initiation / formation of translation preinitiation complex / ribosome disassembly / negative regulation of endoplasmic reticulum unfolded protein response / oxidized pyrimidine DNA binding / response to TNF agonist / positive regulation of base-excision repair / positive regulation of respiratory burst involved in inflammatory response / positive regulation of gastrulation / positive regulation of intrinsic apoptotic signaling pathway in response to DNA damage ...IRES-dependent viral translational initiation / formation of translation preinitiation complex / ribosome disassembly / negative regulation of endoplasmic reticulum unfolded protein response / oxidized pyrimidine DNA binding / response to TNF agonist / positive regulation of base-excision repair / positive regulation of respiratory burst involved in inflammatory response / positive regulation of gastrulation / positive regulation of intrinsic apoptotic signaling pathway in response to DNA damage / protein tyrosine kinase inhibitor activity / positive regulation of ubiquitin-protein transferase activity / IRE1-RACK1-PP2A complex / positive regulation of Golgi to plasma membrane protein transport / nucleolus organization / positive regulation of DNA-templated transcription initiation / TNFR1-mediated ceramide production / negative regulation of RNA splicing / neural crest cell differentiation / ZNF598 and the Ribosome-associated Quality Trigger (RQT) complex dissociate a ribosome stalled on a no-go mRNA / supercoiled DNA binding / negative regulation of DNA repair / PELO:HBS1L and ABCE1 dissociate a ribosome on a non-stop mRNA / cysteine-type endopeptidase activator activity involved in apoptotic process / oxidized purine DNA binding / NF-kappaB complex / cytoplasmic translational initiation / rRNA modification in the nucleus and cytosol / negative regulation of intrinsic apoptotic signaling pathway in response to hydrogen peroxide / erythrocyte homeostasis / regulation of establishment of cell polarity / negative regulation of phagocytosis / negative regulation of bicellular tight junction assembly / ubiquitin-like protein conjugating enzyme binding / cytoplasmic side of rough endoplasmic reticulum membrane / Formation of the ternary complex, and subsequently, the 43S complex / pigmentation / ion channel inhibitor activity / protein kinase A binding / laminin receptor activity / Ribosomal scanning and start codon recognition / positive regulation of mitochondrial depolarization / Translation initiation complex formation / negative regulation of Wnt signaling pathway / fibroblast growth factor binding / Protein hydroxylation / BH3 domain binding / negative regulation of translational frameshifting / TOR signaling / regulation of adenylate cyclase-activating G protein-coupled receptor signaling pathway / monocyte chemotaxis / mTORC1-mediated signalling / SARS-CoV-1 modulates host translation machinery / regulation of cell division / iron-sulfur cluster binding / positive regulation of GTPase activity / cellular response to ethanol / Peptide chain elongation / Selenocysteine synthesis / negative regulation of protein binding / Formation of a pool of free 40S subunits / positive regulation of intrinsic apoptotic signaling pathway by p53 class mediator / Eukaryotic Translation Termination / protein serine/threonine kinase inhibitor activity / negative regulation of respiratory burst involved in inflammatory response / SRP-dependent cotranslational protein targeting to membrane / Response of EIF2AK4 (GCN2) to amino acid deficiency / ubiquitin ligase inhibitor activity / Viral mRNA Translation / endonucleolytic cleavage to generate mature 3'-end of SSU-rRNA from (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / Nonsense Mediated Decay (NMD) independent of the Exon Junction Complex (EJC) / positive regulation of signal transduction by p53 class mediator / negative regulation of ubiquitin-dependent protein catabolic process / GTP hydrolysis and joining of the 60S ribosomal subunit / L13a-mediated translational silencing of Ceruloplasmin expression / Major pathway of rRNA processing in the nucleolus and cytosol / regulation of translational fidelity / positive regulation of microtubule polymerization / phagocytic cup / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / spindle assembly / cellular response to leukemia inhibitory factor / positive regulation of intrinsic apoptotic signaling pathway / Protein methylation / endonucleolytic cleavage in ITS1 to separate SSU-rRNA from 5.8S rRNA and LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / translation regulator activity / Nuclear events stimulated by ALK signaling in cancer / gastrulation / rough endoplasmic reticulum / ribosomal small subunit export from nucleus / Amplification of signal from unattached kinetochores via a MAD2 inhibitory signal / laminin binding / DNA-(apurinic or apyrimidinic site) endonuclease activity / translation initiation factor activity / positive regulation of cell cycle / rescue of stalled cytosolic ribosome / signaling adaptor activity / negative regulation of protein ubiquitination / MDM2/MDM4 family protein binding / Mitotic Prometaphase Similarity search - Function | ||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human)  Hepatitis C virus Hepatitis C virus | ||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 4.2 Å | ||||||
 Authors Authors | Weisser, M. / Schaefer, T. / Leibundgut, M. / Boehringer, D. / Aylett, C.H.S. / Ban, N. | ||||||
 Citation Citation | Journal: Mol Cell / Year: 2017 Title: Structural and Functional Insights into Human Re-initiation Complexes. Authors: Melanie Weisser / Tanja Schäfer / Marc Leibundgut / Daniel Böhringer / Christopher Herbert Stanley Aylett / Nenad Ban /  Abstract: After having translated short upstream open reading frames, ribosomes can re-initiate translation on the same mRNA. This process, referred to as re-initiation, controls the translation of a large ...After having translated short upstream open reading frames, ribosomes can re-initiate translation on the same mRNA. This process, referred to as re-initiation, controls the translation of a large fraction of mammalian cellular mRNAs, many of which are important in cancer. Key ribosomal binding proteins involved in re-initiation are the eukaryotic translation initiation factor 2D (eIF2D) or the homologous complex of MCT-1/DENR. We determined the structures of these factors bound to the human 40S ribosomal subunit in complex with initiator tRNA positioned on an mRNA start codon in the P-site using a combination of cryoelectron microscopy and X-ray crystallography. The structures, supported by biochemical experiments, reveal how eIF2D emulates the function of several canonical translation initiation factors by using three independent, flexibly connected RNA binding domains to simultaneously monitor codon-anticodon interactions in the ribosomal P-site and position the initiator tRNA. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 5oa3.cif.gz | 1.9 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb5oa3.ent.gz | 1.5 MB | Display | PDB format |
| PDBx/mmJSON format | 5oa3.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/oa/5oa3ftp://data.pdbj.org/pub/pdb/validation_reports/oa/5oa3 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  3770MC  5oa9C M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |






-Links
 PDBj
PDBj



































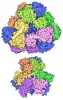








- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-Protein , 3 types, 3 molecules 0fg
| #1: Protein | Mass: 64788.301 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: EIF2D, HCA56, LGTN / Production host:  |
|---|---|
| #36: Protein | Mass: 8453.150 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: Q5RKT7 |
| #37: Protein | Mass: 34857.355 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P63244 |
-RNA chain , 3 types, 3 molecules 123
| #2: RNA chain | Mass: 24238.500 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|---|
| #3: RNA chain | Mass: 602472.688 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: GenBank: 337376 |
| #4: RNA chain | Mass: 88648.414 Da / Num. of mol.: 1 Fragment: (delta domain II mutant, in which domain II of the IRES is replaced with a small stem loop Source method: obtained synthetically Details: HCV IRES mRNA (delta domain II mutant, in which domain II of the IRES is replaced with a small stem loop) Source: (synth.) Hepatitis C virus / References: GenBank: 555439351 |
+40S ribosomal protein ... , 31 types, 31 molecules ABCDEFGHIJKLMNOPQRSTUVWXYZabcde
-Protein/peptide / Non-polymers , 2 types, 4 molecules h
| #38: Protein/peptide | Mass: 3342.255 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) Homo sapiens (human) / References: UniProt: P62945*PLUS |
|---|---|
| #39: Chemical |  Mass: 65.409 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: Zn Mass: 65.409 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: Zn |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Value: 1.4 MDa / Experimental value: NO | ||||||||||||||||||||||||||||||||||||
| Source (natural) |
| ||||||||||||||||||||||||||||||||||||
| Source (recombinant) |
| ||||||||||||||||||||||||||||||||||||
| Buffer solution | pH: 7.6 | ||||||||||||||||||||||||||||||||||||
| Buffer component |
| ||||||||||||||||||||||||||||||||||||
| Specimen | Conc.: 0.11 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||||||||||||||||||
| Specimen support | Grid type: Quantifoil R2/2 | ||||||||||||||||||||||||||||||||||||
| Vitrification | Cryogen name: ETHANE-PROPANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD |
| Image recording | Electron dose: 40 e/Å2 / Film or detector model: FEI FALCON II (4k x 4k) |
- Processing
Processing
| Software | Name: PHENIX / Version: 1.9_1692 / Classification: refinement | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 4.2 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 62177 / Symmetry type: POINT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement | Resolution: 4.2→4.2 Å / SU ML: 0.74 / σ(F): 1.18 / Phase error: 33.96 / Stereochemistry target values: MLHL
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Shrinkage radii: 0.9 Å / VDW probe radii: 1.11 Å / Solvent model: FLAT BULK SOLVENT MODEL | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell |
|
