ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
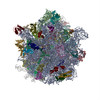
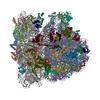
| 登録情報 | データベース: PDB / ID: 3j9y | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
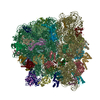
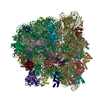
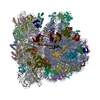
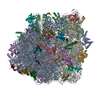
| タイトル | Cryo-EM structure of tetracycline resistance protein TetM bound to a translating E.coli ribosome | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | RIBOSOME / antibiotics / protein synthesis / resistance / TetM / tetracycline / tigecycline / translation | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報ribosome disassembly / negative regulation of cytoplasmic translational initiation / stringent response / mRNA base-pairing translational repressor activity / ornithine decarboxylase inhibitor activity / transcription antitermination factor activity, RNA binding / misfolded RNA binding / Group I intron splicing / RNA folding / transcriptional attenuation ...ribosome disassembly / negative regulation of cytoplasmic translational initiation / stringent response / mRNA base-pairing translational repressor activity / ornithine decarboxylase inhibitor activity / transcription antitermination factor activity, RNA binding / misfolded RNA binding / Group I intron splicing / RNA folding / transcriptional attenuation / endoribonuclease inhibitor activity / RNA-binding transcription regulator activity / positive regulation of ribosome biogenesis / negative regulation of cytoplasmic translation / translational termination / four-way junction DNA binding / DnaA-L2 complex / translation repressor activity / negative regulation of translational initiation / negative regulation of DNA-templated DNA replication initiation / regulation of mRNA stability / translational initiation / ribosome assembly / mRNA regulatory element binding translation repressor activity / positive regulation of RNA splicing / assembly of large subunit precursor of preribosome / transcription elongation factor complex / DNA endonuclease activity / regulation of DNA-templated transcription elongation / cytosolic ribosome assembly / transcription antitermination / response to reactive oxygen species / regulation of cell growth / DNA-templated transcription termination / maintenance of translational fidelity / response to radiation / ribosomal large subunit assembly / mRNA 5'-UTR binding / large ribosomal subunit / ribosome biogenesis / ribosome binding / regulation of translation / ribosomal small subunit biogenesis / ribosomal small subunit assembly / small ribosomal subunit / small ribosomal subunit rRNA binding / transferase activity / 5S rRNA binding / large ribosomal subunit rRNA binding / cytosolic small ribosomal subunit / cytosolic large ribosomal subunit / tRNA binding / molecular adaptor activity / cytoplasmic translation / rRNA binding / negative regulation of translation / ribosome / structural constituent of ribosome / translation / response to antibiotic / GTPase activity / negative regulation of DNA-templated transcription / mRNA binding / GTP binding / DNA binding / RNA binding / zinc ion binding / membrane / cytoplasm / cytosol 類似検索 - 分子機能 | |||||||||
| 生物種 |   Enterococcus faecalis (乳酸球菌) Enterococcus faecalis (乳酸球菌) | |||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.9 Å | |||||||||
 データ登録者 データ登録者 | Arenz, S. / Nguyen, F. / Beckmann, R. / Wilson, D.N. | |||||||||
 引用 引用 | ジャーナル: Proc Natl Acad Sci U S A / 年: 2015 タイトル: Cryo-EM structure of the tetracycline resistance protein TetM in complex with a translating ribosome at 3.9-Å resolution. 著者: Stefan Arenz / Fabian Nguyen / Roland Beckmann / Daniel N Wilson /  要旨: Ribosome protection proteins (RPPs) confer resistance to tetracycline by binding to the ribosome and chasing the drug from its binding site. Current models for RPP action are derived from 7.2- to 16- ...Ribosome protection proteins (RPPs) confer resistance to tetracycline by binding to the ribosome and chasing the drug from its binding site. Current models for RPP action are derived from 7.2- to 16-Å resolution structures of RPPs bound to vacant or nontranslating ribosomes. Here we present a cryo-electron microscopy reconstruction of the RPP TetM in complex with a translating ribosome at 3.9-Å resolution. The structure reveals the contacts of TetM with the ribosome, including interaction between the conserved and functionally critical C-terminal extension of TetM with a unique splayed conformation of nucleotides A1492 and A1493 at the decoding center of the small subunit. The resolution enables us to unambiguously model the side chains of the amino acid residues comprising loop III in domain IV of TetM, revealing that the tyrosine residues Y506 and Y507 are not responsible for drug-release as suggested previously but rather for intrafactor contacts that appear to stabilize the conformation of loop III. Instead, Pro509 at the tip of loop III is located directly within the tetracycline binding site where it interacts with nucleotide C1054 of the 16S rRNA, such that RPP action uses Pro509, rather than Y506/Y507, to directly dislodge and release tetracycline from the ribosome. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 3j9y.cif.gz | 3.7 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb3j9y.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 3j9y.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 3j9y_validation.pdf.gz | 1.5 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 3j9y_full_validation.pdf.gz | 1.6 MB | 表示 | |
| XML形式データ | 3j9y_validation.xml.gz | 221.7 KB | 表示 | |
| CIF形式データ | 3j9y_validation.cif.gz | 382.2 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/j9/3j9yftp://data.pdbj.org/pub/pdb/validation_reports/j9/3j9y | HTTPS FTP |
-関連構造データ



-リンク
 PDBj
PDBj





























- 集合体
集合体
| 登録構造単位 | 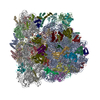
|
|---|---|
| 1 |
|
-要素
-RNA鎖 , 5種, 5分子 avxAB
| #1: RNA鎖 | 分子量: 498909.844 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #15: RNA鎖 | 分子量: 24978.098 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #16: RNA鎖 | 分子量: 3492.122 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #25: RNA鎖 | 分子量: 941521.375 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #26: RNA鎖 | 分子量: 38813.133 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-30S ribosomal protein ... , 20種, 20分子 bdefhklopqrtucgijmns
| #2: タンパク質 | 分子量: 26652.557 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #3: タンパク質 | 分子量: 23514.199 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #4: タンパク質 | 分子量: 17629.398 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #5: タンパク質 | 分子量: 15727.512 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #6: タンパク質 | 分子量: 14146.557 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #7: タンパク質 | 分子量: 13870.975 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #8: タンパク質 | 分子量: 13768.157 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #9: タンパク質 | 分子量: 10290.816 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #10: タンパク質 | 分子量: 9207.572 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #11: タンパク質 | 分子量: 9724.491 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #12: タンパク質 | 分子量: 9005.472 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #13: タンパク質 | 分子量: 9708.464 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #14: タンパク質 | 分子量: 8524.039 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #18: タンパク質 | 分子量: 26031.316 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #19: タンパク質 | 分子量: 20055.156 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #20: タンパク質 | 分子量: 14886.270 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #21: タンパク質 | 分子量: 11755.597 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #22: タンパク質 | 分子量: 13128.467 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #23: タンパク質 | 分子量: 11677.637 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #24: タンパク質 | 分子量: 10455.355 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-タンパク質 , 1種, 1分子 w
| #17: タンパク質 | 分子量: 72542.883 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Enterococcus faecalis (乳酸球菌) / 発現宿主: |
|---|
+50S ribosomal protein ... , 32種, 32分子 CDEFGHIJKLMNOPQRSTUVWXYZ01234567
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 緩衝液 | 名称: 50 mM HEPES-KOH, 100 mM KOAc, 25 mM Mg(OAc)2, 6 mM b-mercaptoethanol pH: 7.4 詳細: 50 mM HEPES-KOH, 100 mM KOAc, 25 mM Mg(OAc)2, 6 mM b-mercaptoethanol | ||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 詳細: Plunged into liquid ethane (FEI VITROBOT MARK IV) |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS / 日付: 2014年3月14日 |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: SPOT SCAN FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: SPOT SCAN |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 125085 X / 最大 デフォーカス(公称値): 3500 nm / 最小 デフォーカス(公称値): 700 nm / Cs: 0 mm |
| 試料ホルダ | 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 電子線照射量: 28 e/Å2 フィルム・検出器のモデル: FEI FALCON II (4k x 4k) |
| 画像スキャン | デジタル画像の数: 2753 |
| 放射 | プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray |
| 放射波長 | 相対比: 1 |
- 解析
解析
| EMソフトウェア |
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | 詳細: Defocus groups | ||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||
| 3次元再構成 | 解像度: 3.9 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 78186 / ピクセルサイズ(公称値): 1.108 Å / ピクセルサイズ(実測値): 1.108 Å 詳細: Since images from microscopy were processed in the absence of spatial frequencies higher than 8 A, an FSC cut-off value of 0.143 was used for average resolution determination of 3.9 A ...詳細: Since images from microscopy were processed in the absence of spatial frequencies higher than 8 A, an FSC cut-off value of 0.143 was used for average resolution determination of 3.9 A (Scheres and Chen, 2012). The final map was sharpened by applying an automatically determined negative B-factor using the program EMBFACTOR (Fernandez et al, 2008). (Single particle--Applied symmetry: C1) 対称性のタイプ: POINT | ||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT / 空間: REAL / 詳細: REFINEMENT PROTOCOL--rigid body | ||||||||||||
| 原子モデル構築 | PDB-ID: 5AFI | ||||||||||||
| 精密化ステップ | サイクル: LAST
|