 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 3zko | ||||||
|---|---|---|---|---|---|---|---|
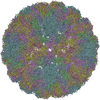
| Title | The structure of ''breathing'' dengue virus. | ||||||
 Components Components | ENVELOPE GLYCOPROTEIN | ||||||
 Keywords Keywords |  VIRUS / BREATHING STRUCTURE / DYNAMIC STRUCTURE VIRUS / BREATHING STRUCTURE / DYNAMIC STRUCTURE | ||||||
| Function / homology |  Function and homology information Function and homology informationsymbiont-mediated suppression of host JAK-STAT cascade via inhibition of host TYK2 activity / flavivirin / host cell mitochondrion / symbiont-mediated suppression of host JAK-STAT cascade via inhibition of STAT2 activity / symbiont-mediated suppression of host cytoplasmic pattern recognition receptor signaling pathway via inhibition of MAVS activity / viral capsid / : / nucleoside-triphosphate phosphatase / double-stranded RNA binding / protein complex oligomerization ...symbiont-mediated suppression of host JAK-STAT cascade via inhibition of host TYK2 activity / flavivirin / host cell mitochondrion / symbiont-mediated suppression of host JAK-STAT cascade via inhibition of STAT2 activity / symbiont-mediated suppression of host cytoplasmic pattern recognition receptor signaling pathway via inhibition of MAVS activity / viral capsid / : / nucleoside-triphosphate phosphatase / double-stranded RNA binding / protein complex oligomerization / monoatomic ion channel activity / mRNA (guanine-N7)-methyltransferase / methyltransferase cap1 / clathrin-dependent endocytosis of virus by host cell / mRNA (nucleoside-2'-O-)-methyltransferase activity / mRNA 5'-cap (guanine-N7-)-methyltransferase activity / RNA helicase activity / host cell endoplasmic reticulum membrane / host cell perinuclear region of cytoplasm / protein dimerization activity / RNA helicase / induction by virus of host autophagy / RNA-directed RNA polymerase / viral RNA genome replication / RNA-dependent RNA polymerase activity / serine-type endopeptidase activity / fusion of virus membrane with host endosome membrane / viral envelope / symbiont-mediated suppression of host type I interferon-mediated signaling pathway / host cell nucleus / virion attachment to host cell / virion membrane / structural molecule activity / ATP hydrolysis activity / proteolysis / extracellular region / ATP binding / membrane / metal ion bindingSimilarity search - Function | ||||||
| Biological species |  DENGUE VIRUS 2 DENGUE VIRUS 2 | ||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 13.7 Å | ||||||
| Model type details | CA ATOMS ONLY, CHAIN A, B, C | ||||||
 Authors Authors | Fibriansah, G. / Ng, T.S. / Kostyuchenko, V.A. / Lee, S. / Wang, J. / Lok, S.M. | ||||||
 Citation Citation | Journal: J Virol / Year: 2013 Title: Structural changes in dengue virus when exposed to a temperature of 37°C. Authors: Guntur Fibriansah / Thiam-Seng Ng / Victor A Kostyuchenko / Jaime Lee / Sumarlin Lee / Jiaqi Wang / Shee-Mei Lok /  Abstract: Previous binding studies of antibodies that recognized a partially or fully hidden epitope suggest that insect cell-derived dengue virus undergoes structural changes at an elevated temperature. This ...Previous binding studies of antibodies that recognized a partially or fully hidden epitope suggest that insect cell-derived dengue virus undergoes structural changes at an elevated temperature. This was confirmed by our cryo-electron microscopy images of dengue virus incubated at 37°C, where viruses change their surface from smooth to rough. Here we present the cryo-electron microscopy structures of dengue virus at 37°C. Image analysis showed four classes of particles. The three-dimensional (3D) map of one of these classes, representing half of the imaged virus population, shows that the E protein shell has expanded and there is a hole at the 3-fold vertices. Fitting E protein structures into the map suggests that all of the interdimeric and some intradimeric E protein interactions are weakened. The accessibility of some previously found cryptic epitopes on this class of particles is discussed. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 3zko.cif.gz | 50.7 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb3zko.ent.gz | 33 KB | Display | PDB format |
| PDBx/mmJSON format | 3zko.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/zk/3zkoftp://data.pdbj.org/pub/pdb/validation_reports/zk/3zko | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  2296MC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |







-Links
 PDBj
PDBj
- Assembly
Assembly
| Deposited unit | 
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | x 60
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3 | x 5
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4 | x 6
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5 | 
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: (Schoenflies symbol: I (icosahedral)) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Noncrystallographic symmetry (NCS) | NCS oper:
|