 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7z13 | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
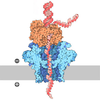
| タイトル | S. cerevisiae CMGE dimer nucleating origin DNA melting | |||||||||||||||||||||||||||||||||
 要素 要素 |
| |||||||||||||||||||||||||||||||||
 キーワード キーワード |  REPLICATION (DNA複製) / DNA replication (DNA複製) / helicase (ヘリカーゼ) / initiation / DNA origin REPLICATION (DNA複製) / DNA replication (DNA複製) / helicase (ヘリカーゼ) / initiation / DNA origin | |||||||||||||||||||||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報DNA-templated DNA replication maintenance of fidelity / 遺伝子変換 / Unwinding of DNA / DNA replication initiation / epsilon DNA polymerase complex / MCM core complex / Assembly of the pre-replicative complex / Switching of origins to a post-replicative state / nuclear DNA replication / MCM complex binding ...DNA-templated DNA replication maintenance of fidelity / 遺伝子変換 / Unwinding of DNA / DNA replication initiation / epsilon DNA polymerase complex / MCM core complex / Assembly of the pre-replicative complex / Switching of origins to a post-replicative state / nuclear DNA replication / MCM complex binding / GINS complex / mitotic DNA replication preinitiation complex assembly / premeiotic DNA replication / pre-replicative complex assembly involved in nuclear cell cycle DNA replication / nucleotide-excision repair, DNA gap filling / SUMO binding / mitotic DNA replication / DNA replication proofreading / Activation of the pre-replicative complex / CMG complex / single-stranded DNA 3'-5' DNA exonuclease activity / nuclear pre-replicative complex / Activation of ATR in response to replication stress / MCM complex / DNA replication preinitiation complex / double-strand break repair via break-induced replication / mitotic DNA replication checkpoint signaling / replication fork protection complex / mitotic DNA replication initiation / single-stranded DNA helicase activity / mitotic intra-S DNA damage checkpoint signaling / 加水分解酵素; エステル加水分解酵素; 5'-リン酸モノエステル産生エンドデオキシリボヌクレアーゼ / silent mating-type cassette heterochromatin formation / DNA strand elongation involved in DNA replication / mitotic sister chromatid cohesion / leading strand elongation / nuclear chromosome / DNA unwinding involved in DNA replication / DNA replication origin binding / nuclear replication fork / subtelomeric heterochromatin formation / DNA replication initiation / error-prone translesion synthesis / base-excision repair, gap-filling / DNA helicase activity / DNA複製 / helicase activity / デオキシリボ核酸 / 塩基除去修復 / DNA-templated DNA replication / double-strand break repair via nonhomologous end joining / double-strand break repair / single-stranded DNA binding / mitotic cell cycle / 4 iron, 4 sulfur cluster binding / double-stranded DNA binding / ヘリカーゼ / DNA複製 / chromosome, telomeric region / DNAポリメラーゼ / DNA-directed DNA polymerase activity / hydrolase activity / 細胞周期 / nucleotide binding / mRNA binding / chromatin binding / ATP hydrolysis activity / DNA binding / zinc ion binding / 核質 / ATP binding / metal ion binding / 細胞核 / 細胞質類似検索 - 分子機能 | |||||||||||||||||||||||||||||||||
| 生物種 |  Saccharomyces cerevisiae (パン酵母)DNA molecule (デオキシリボ核酸) Saccharomyces cerevisiae (パン酵母)DNA molecule (デオキシリボ核酸) | |||||||||||||||||||||||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.4 Å | |||||||||||||||||||||||||||||||||
 データ登録者 データ登録者 | Lewis, J.S. / Sousa, J.S. / Costa, A. | |||||||||||||||||||||||||||||||||
| 資金援助 | European Union,  フランス, フランス,  英国, 10件 英国, 10件
| |||||||||||||||||||||||||||||||||
 引用 引用 | ジャーナル: Nature / 年: 2022 タイトル: Mechanism of replication origin melting nucleated by CMG helicase assembly. 著者: Jacob S Lewis / Marta H Gross / Joana Sousa / Sarah S Henrikus / Julia F Greiwe / Andrea Nans / John F X Diffley / Alessandro Costa / 要旨: The activation of eukaryotic origins of replication occurs in temporally separated steps to ensure that chromosomes are copied only once per cell cycle. First, the MCM helicase is loaded onto duplex ...The activation of eukaryotic origins of replication occurs in temporally separated steps to ensure that chromosomes are copied only once per cell cycle. First, the MCM helicase is loaded onto duplex DNA as an inactive double hexamer. Activation occurs after the recruitment of a set of firing factors that assemble two Cdc45-MCM-GINS (CMG) holo-helicases. CMG formation leads to the underwinding of DNA on the path to the establishment of the replication fork, but whether DNA becomes melted at this stage is unknown. Here we use cryo-electron microscopy to image ATP-dependent CMG assembly on a chromatinized origin, reconstituted in vitro with purified yeast proteins. We find that CMG formation disrupts the double hexamer interface and thereby exposes duplex DNA in between the two CMGs. The two helicases remain tethered, which gives rise to a splayed dimer, with implications for origin activation and replisome integrity. Inside each MCM ring, the double helix becomes untwisted and base pairing is broken. This comes as the result of ATP-triggered conformational changes in MCM that involve DNA stretching and protein-mediated stabilization of three orphan bases. Mcm2 pore-loop residues that engage DNA in our structure are dispensable for double hexamer loading and CMG formation, but are essential to untwist the DNA and promote replication. Our results explain how ATP binding nucleates origin DNA melting by the CMG and maintains replisome stability at initiation. | |||||||||||||||||||||||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7z13.cif.gz | 2.4 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7z13.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 7z13.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/z1/7z13ftp://data.pdbj.org/pub/pdb/validation_reports/z1/7z13 | HTTPS FTP |
|---|
-関連構造データ



-リンク
 PDBj
PDBj







































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-DNA replication licensing factor ... , 5種, 10分子 2a3b4c6e7f
| #1: タンパク質 | 分子量: 98911.539 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae (パン酵母)遺伝子: PACBIOSEQ_LOCUS187, PACBIOSEQ_LOCUS193, PACBIOSEQ_LOCUS195, PACBIOSEQ_LOCUS196, SCNYR20_0007007400, SCP684_0007007100 発現宿主: Saccharomyces cerevisiae (パン酵母) / 参照: UniProt: A0A6A5Q1S9, ヘリカーゼ#2: タンパク質 | 分子量: 111720.242 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae (パン酵母)株: ATCC 204508 / S288c / 遺伝子: MCM3, YEL032W, SYGP-ORF23 / 発現宿主: Saccharomyces cerevisiae (パン酵母) / 参照: UniProt: P24279, ヘリカーゼ#3: タンパク質 | 分子量: 105138.375 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae (パン酵母)株: ATCC 204508 / S288c / 遺伝子: MCM4, CDC54, HCD21, YPR019W, YP9531.13 / 発現宿主: Saccharomyces cerevisiae (パン酵母) / 参照: UniProt: P30665, ヘリカーゼ#5: タンパク質 | 分子量: 113110.211 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae (パン酵母)株: ATCC 204508 / S288c / 遺伝子: MCM6, YGL201C / 発現宿主: Saccharomyces cerevisiae (パン酵母) / 参照: UniProt: P53091, ヘリカーゼ#6: タンパク質 | 分子量: 95049.875 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae (パン酵母)遺伝子: MCM7, PACBIOSEQ_LOCUS429 / 発現宿主: Saccharomyces cerevisiae (パン酵母) / 参照: UniProt: A0A6A5Q4N0, ヘリカーゼ |
|---|
-タンパク質 , 2種, 4分子 5dEL
| #4: タンパク質 | ヘリカーゼ 分子量: 86505.734 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae (パン酵母)遺伝子: PACBIOSEQ_LOCUS4054, PACBIOSEQ_LOCUS4112, PACBIOSEQ_LOCUS4129, PACBIOSEQ_LOCUS4153, PACBIOSEQ_LOCUS4202, SCNYR20_0004029000, SCP684_0004028600 発現宿主: Saccharomyces cerevisiae (パン酵母) / 参照: UniProt: A0A6A5PUY8, ヘリカーゼ#11: タンパク質 | 分子量: 75154.703 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae (パン酵母)株: ATCC 204508 / S288c / 遺伝子: CDC45, SLD4, YLR103C, L8004.11 / 発現宿主: Saccharomyces cerevisiae (パン酵母) / 参照: UniProt: Q08032 |
|---|
-DNA鎖 , 2種, 2分子 AB
| #7: DNA鎖 | 分子量: 16320.634 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) DNA molecule (デオキシリボ核酸) |
|---|---|
| #8: DNA鎖 | 分子量: 16311.620 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) DNA molecule (デオキシリボ核酸) |
-DNA replication complex GINS protein ... , 4種, 8分子 CJDKHOIP
| #9: タンパク質 | 分子量: 25718.070 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae (パン酵母)株: ATCC 204508 / S288c / 遺伝子: PSF3, YOL146W / 発現宿主:  Escherichia coli (大腸菌) / 参照: UniProt: Q12146 Escherichia coli (大腸菌) / 参照: UniProt: Q12146#10: タンパク質 | 分子量: 33983.617 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae (パン酵母)株: ATCC 204508 / S288c / 遺伝子: SLD5, YDR489W / 発現宿主: Escherichia coli (大腸菌) / 参照: UniProt: Q03406#13: タンパク質 | 分子量: 24230.576 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae (パン酵母)遺伝子: PACBIOSEQ_LOCUS944, PACBIOSEQ_LOCUS956, PACBIOSEQ_LOCUS958, SCNYR20_0001022500, SCP684_0001022000 発現宿主: Escherichia coli (大腸菌) / 参照: UniProt: A0A6A5Q203#14: タンパク質 | 分子量: 25096.807 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae (パン酵母)遺伝子: PACBIOSEQ_LOCUS3163, PACBIOSEQ_LOCUS3191, PACBIOSEQ_LOCUS3224, PACBIOSEQ_LOCUS3231, PACBIOSEQ_LOCUS3255, SCNYR20_0009012300, SCP684_0009011800 発現宿主: Escherichia coli (大腸菌) / 参照: UniProt: A0A6A5PX40 |
|---|
-DNA polymerase epsilon ... , 2種, 4分子 FMNQ
| #12: タンパク質 | / DNA polymerase II subunit 2 分子量: 78425.852 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae (パン酵母)株: ATCC 204508 / S288c / 遺伝子: DPB2, YPR175W, P9705.7 / 発現宿主: Saccharomyces cerevisiae (パン酵母) / 参照: UniProt: P24482#15: タンパク質 | 分子量: 255992.484 Da / 分子数: 2 / 由来タイプ: 組換発現 由来: (組換発現) Saccharomyces cerevisiae (パン酵母)株: ATCC 204508 / S288c / 遺伝子: POL2, DUN2, YNL262W, N0825 / 発現宿主: Saccharomyces cerevisiae (パン酵母)参照: UniProt: P21951, DNAポリメラーゼ, 加水分解酵素; エステル加水分解酵素; 5'-リン酸モノエステル産生エンドデオキシリボヌクレアーゼ |
|---|
-非ポリマー , 4種, 32分子 



| #16: 化合物 | ChemComp-ATP / アデノシン三リン酸 分子量: 507.181 Da / 分子数: 8 / 由来タイプ: 合成 / 式: C10H16N5O13P3 / タイプ: SUBJECT OF INVESTIGATION / コメント: ATP, エネルギー貯蔵分子*YM 分子量: 507.181 Da / 分子数: 8 / 由来タイプ: 合成 / 式: C10H16N5O13P3 / タイプ: SUBJECT OF INVESTIGATION / コメント: ATP, エネルギー貯蔵分子*YM#17: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 14 / 由来タイプ: 合成 / 式: Zn / タイプ: SUBJECT OF INVESTIGATION 分子量: 65.409 Da / 分子数: 14 / 由来タイプ: 合成 / 式: Zn / タイプ: SUBJECT OF INVESTIGATION#18: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 6 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION 分子量: 24.305 Da / 分子数: 6 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION#19: 化合物 | ChemComp-ADP / アデノシン二リン酸 分子量: 427.201 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / タイプ: SUBJECT OF INVESTIGATION / コメント: ADP, エネルギー貯蔵分子*YM 分子量: 427.201 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C10H15N5O10P2 / タイプ: SUBJECT OF INVESTIGATION / コメント: ADP, エネルギー貯蔵分子*YM |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: S. cerevisiae CMGE dimer nucleating origin DNA melting タイプ: COMPLEX 詳細: Dimeric model reconstituted from symmetry expanded monomer (PDB entry 7QHS). Entity ID: #1-#15 / 由来: RECOMBINANT |
|---|---|
| 分子量 | 実験値: NO |
| 由来(天然) | 生物種: Saccharomyces cerevisiae (パン酵母) |
| 由来(組換発現) | 生物種: Saccharomyces cerevisiae (パン酵母) |
| 緩衝液 | pH: 7.5 |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES 詳細: four microlitres of sample was applied on a grid and incubated for 2 min at room temperature before blotting with filter paper for 5.5 s and plunge-freezing in liquid ethane. |
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源: FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: OTHER |
| 電子レンズ | モード: BRIGHT FIELDBright-field microscopy / 倍率(公称値): 130000 X / 最大 デフォーカス(公称値): 4400 nm / 最小 デフォーカス(公称値): 2000 nm / Cs: 2.7 mm / C2レンズ絞り径: 50 µm |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 平均露光時間: 10 sec. / 電子線照射量: 1.6 e/Å2 / 検出モード: COUNTING フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 撮影したグリッド数: 2 / 実像数: 65286 |
| 画像スキャン | 動画フレーム数/画像: 32 |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.4 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 71348 / 対称性のタイプ: POINT | ||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT 詳細: One additional base pair has been built to connect the DNA molecules from the two individual symmetry expanded monomers. | ||||||||||||||||||||||||
| 原子モデル構築 |
|
