 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 1fbk | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | CRYSTAL STRUCTURE OF CYTOPLASMICALLY OPEN CONFORMATION OF BACTERIORHODOPSIN | ||||||
 要素 要素 | BACTERIORHODOPSIN バクテリオロドプシン バクテリオロドプシン | ||||||
 キーワード キーワード | PROTON TRANSPORT (プロトンポンプ) / PROTON PUMP (プロトンポンプ) / MEMBRANE PROTEIN (膜タンパク質) / RETINAL PROTEIN (レチナール) / TWO-DIMENSIONAL CRYSTAL ELECTRON DIFFRACTION / SINGLE CRYSTAL (単結晶) | ||||||
| 機能・相同性 |  機能・相同性情報photoreceptor activity / phototransduction / proton transmembrane transport / monoatomic ion channel activity / 細胞膜 機能・相同性情報photoreceptor activity / phototransduction / proton transmembrane transport / monoatomic ion channel activity / 細胞膜類似検索 - 分子機能 | ||||||
| 生物種 |  Halobacterium salinarum (好塩性) Halobacterium salinarum (好塩性) | ||||||
| 手法 | 電子線結晶学 / クライオ電子顕微鏡法 / 解像度: 3.2 Å | ||||||
 データ登録者 データ登録者 | Subramaniam, S. / Henderson, R. | ||||||
 引用 引用 | ジャーナル: Nature / 年: 2000 タイトル: Molecular mechanism of vectorial proton translocation by bacteriorhodopsin. 著者: S Subramaniam / R Henderson /  要旨: Bacteriorhodopsin, a membrane protein with a relative molecular mass of 27,000, is a light driven pump which transports protons across the cell membrane of the halophilic organism Halobacterium ...Bacteriorhodopsin, a membrane protein with a relative molecular mass of 27,000, is a light driven pump which transports protons across the cell membrane of the halophilic organism Halobacterium salinarum. The chromophore retinal is covalently attached to the protein via a protonated Schiff base. Upon illumination, retinal is isomerized. The Schiff base then releases a proton to the extracellular medium, and is subsequently reprotonated from the cytoplasm. An atomic model for bacteriorhodopsin was first determined by Henderson et al, and has been confirmed and extended by work in a number of laboratories in the last few years. Here we present an atomic model for structural changes involved in the vectorial, light-driven transport of protons by bacteriorhodopsin. A 'switch' mechanism ensures the vectorial nature of pumping. First, retinal unbends, triggered by loss of the Schiff base proton, and second, a protein conformational change occurs. This conformational change, which we have determined by electron crystallography at atomic (3.2 A in-plane and 3.6 A vertical) resolution, is largely localized to helices F and G, and provides an 'opening' of the protein to protons on the cytoplasmic side of the membrane. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 1fbk.cif.gz | 54.6 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb1fbk.ent.gz | 41.3 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 1fbk.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/fb/1fbkftp://data.pdbj.org/pub/pdb/validation_reports/fb/1fbk | HTTPS FTP |
|---|
-関連構造データ







-リンク
 PDBj
PDBj



- 集合体
集合体
| 登録構造単位 | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||
| 単位格子 |
| ||||||||
| 詳細 | Trimer is present in vivo. The crystals contain trimers related by crystal symmetry P3. |
-要素
| #1: タンパク質 | バクテリオロドプシン 分子量: 26678.330 Da / 分子数: 1 / 変異: D96G,F171C,F219L / 由来タイプ: 天然 / 由来: (天然) Halobacterium salinarum (好塩性) / 参照: UniProt: P02945 |
|---|---|
| #2: 化合物 | ChemComp-RET / レチナール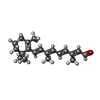  分子量: 284.436 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C20H28O 分子量: 284.436 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C20H28O |
-実験情報
-実験
| 実験 | 手法: 電子線結晶学 / 使用した結晶の数: 402 |
|---|---|
| EM実験 | 試料の集合状態: 2D ARRAY / 3次元再構成法: 電子線結晶学 |
| 結晶の対称性 | ∠γ: 120 ° / A: 62.45 Å / B: 62.45 Å / C: 100.9 Å / Space group name H-M: P3 |
- 試料調製
試料調製
| 構成要素 | 名称: CYTOPLASMICALLY OPEN CONFORMATION OF BACTERIORHODOPSIN 由来: RECOMBINANT | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: .027 MDa / 実験値: NO | ||||||||||||||||||||||||||||||
| 由来(天然) | 生物種: Halobacterium salinarum (好塩性) | ||||||||||||||||||||||||||||||
| 試料 | 包埋: YES / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||
| EM embedding | Material: glucose or trehalose | ||||||||||||||||||||||||||||||
| 結晶化 | 温度: 310 K / 手法: naturally occurring in vivo / pH: 7 詳細: crystals are increased in size by fusion and annealing using detergents, pH 7, naturally occurring in vivo, temperature 37K | ||||||||||||||||||||||||||||||
| 結晶化 | *PLUS 温度: 4 ℃ / pH: 5.6 / 手法: unknown | ||||||||||||||||||||||||||||||
| 溶液の組成 | *PLUS
|
-データ収集
| 顕微鏡 | モデル: FEI/PHILIPS CM12 / 詳細: imaging date 1998-12-01 |
|---|---|
| 電子銃 | 照射モード: FLOOD BEAM |
| 電子レンズ | モード: DIFFRACTION回折 |
| 撮影 | フィルム・検出器のモデル: GENERIC CCD / Num. of diffraction images: 1000 |
| 回折 | 平均測定温度: 93 K |
| 放射光源 | 由来: ELECTRON MICROSCOPE / タイプ: その他 / 波長: 0.033 |
| 検出器 | タイプ: その他 / 検出器: CCD / 日付: 1998年12月1日 |
| 放射 | プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: electron |
| 放射波長 | 波長: 0.033 Å / 相対比: 1 |
| 反射 | 解像度: 3.2→200 Å / Num. all: 7298 / Num. obs: 5668 / % possible obs: 77.7 % / Observed criterion σ(I): 0 / Rmerge(I) obs: 0.173 |
- 解析
解析
| ソフトウェア | 名称: CNS / バージョン: 0.9 / 分類: 精密化 | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 結晶の対称性 | ∠γ: 120 ° / A: 62.45 Å / B: 62.45 Å / C: 100.9 Å / Space group name H-M: P3 | |||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.2 Å / 解像度の算出法: DIFFRACTION PATTERN/LAYERLINES 詳細: 402 patterns merged (merging R 17.3%) to generate set of lattice lines covering ~87% of reciprocal space, with resolution of 3.2 A in-plane and 3.6 A vertically [primary citation] | |||||||||||||||||||||||||
| 精密化 | 解像度: 3.2→200 Å / 立体化学のターゲット値: Engh & Huber 詳細: Each diffraction pattern was automatically indexed, and the spot intensities were integrated either using a raster (for patterns recorded at specimen tilts less than 30 degrees) or using ...詳細: Each diffraction pattern was automatically indexed, and the spot intensities were integrated either using a raster (for patterns recorded at specimen tilts less than 30 degrees) or using profile fitting (for patterns recorded at specimen tilts at or greater than 30 degrees, which represented about 80% of the total data set). Each pattern was then compared to the curves recorded for wild-type bacteriorhodopsin in glucose at -100 degrees C [Ceska and Henderson J. Mol. Biol. 213: 539-560 (1990)], and the relative proportions of the four different twins determined. This exercise was carried out with all four theoretically possible orientations of the crystal axes relative to the previous reference curves to ensure that the data were merged correctly. From the initial set of 486 patterns chosen, 286 minimally twinned diffraction patterns were selected in which the major twin proportion was greater than 0.8. These 286 patterns were merged using the wild-type lattice lines as a reference and lattice lines were fitted to the data to obtain an initial approximately merged set of lattice lines (merge #1) describing the structure of the triple mutant. The original set of 486 patterns was then merged against the new lattice curves to redetermine the twin proportions more accurately. The merging parameters for each crystal were inspected carefully again, and 84 crystals for which the major twin proportion was less than 0.70 were excluded from the data set. The remaining 402 substantially untwinned diffraction patterns were used to generate a new set of curves and the procedure repeated to create a stable set of lattice lines. The merged data were further improved by using an estimate of sigma values for each reflection, and by the inclusion of an individual weighting factor for each diffraction pattern using procedures described by Grigorieff and Henderson [Ultramicroscopy 60: 295-309 (1995)]. Two cycles of this refinement were carried out to obtain a final set of merged curves. The curves were sampled at 1/100 Angstroms (approximately twice the thickness of the membrane) to obtain a set of intensities at H,K,L values so that the data could be further processed with standard X-ray crystallographic programs. For the tilt angles used, the maximal possible theoretical completeness of the data set is ~87%. The completeness of our data is close to this limit up to 3.5 Angstroms. The completeness drops to 77.7 % when all of the data to 3.2 Angstroms is included.
| |||||||||||||||||||||||||
| 精密化ステップ | サイクル: LAST / 解像度: 3.2→200 Å
| |||||||||||||||||||||||||
| 拘束条件 |
|
