 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 3lue | ||||||
|---|---|---|---|---|---|---|---|
| Title | Model of alpha-actinin CH1 bound to F-actin | ||||||
 Components Components |
| ||||||
 Keywords Keywords | STRUCTURAL PROTEIN / calponin homology domains / Acetylation / ATP-binding / Cytoplasm / Cytoskeleton / Methylation / Nucleotide-binding / Phosphoprotein / Actin-binding / Calcium / Polymorphism / Deafness / Disease mutation / Dystonia | ||||||
| Function / homology |  Function and homology information Function and homology informationpositive regulation of glucose catabolic process to lactate via pyruvate / negative regulation of relaxation of muscle / regulation of the force of skeletal muscle contraction / skeletal muscle atrophy / positive regulation of skeletal muscle fiber development / positive regulation of skeletal muscle tissue growth / response to denervation involved in regulation of muscle adaptation / positive regulation of norepinephrine uptake / cellular response to cytochalasin B / positive regulation of fast-twitch skeletal muscle fiber contraction ...positive regulation of glucose catabolic process to lactate via pyruvate / negative regulation of relaxation of muscle / regulation of the force of skeletal muscle contraction / skeletal muscle atrophy / positive regulation of skeletal muscle fiber development / positive regulation of skeletal muscle tissue growth / response to denervation involved in regulation of muscle adaptation / positive regulation of norepinephrine uptake / cellular response to cytochalasin B / positive regulation of fast-twitch skeletal muscle fiber contraction / regulation of transepithelial transport / morphogenesis of a polarized epithelium / bBAF complex / postsynaptic actin cytoskeleton organization / npBAF complex / nBAF complex / protein localization to adherens junction / brahma complex / postsynaptic actin cytoskeleton / Tat protein binding / structural constituent of postsynaptic actin cytoskeleton / transition between fast and slow fiber / positive regulation of bone mineralization involved in bone maturation / GBAF complex / dense body / Formation of annular gap junctions / regulation of G0 to G1 transition / Gap junction degradation / Cell-extracellular matrix interactions / Folding of actin by CCT/TriC / apical protein localization / muscle cell development / regulation of double-strand break repair / adherens junction assembly / regulation of nucleotide-excision repair / Prefoldin mediated transfer of substrate to CCT/TriC / RSC-type complex / negative regulation of oxidative phosphorylation / focal adhesion assembly / RHOF GTPase cycle / Regulation of MITF-M-dependent genes involved in pigmentation / Adherens junctions interactions / Striated Muscle Contraction / tight junction / bone morphogenesis / negative regulation of glycolytic process / Nephrin family interactions / Sensory processing of sound by outer hair cells of the cochlea / regulation of norepinephrine uptake / Interaction between L1 and Ankyrins / Sensory processing of sound by inner hair cells of the cochlea / regulation of mitotic metaphase/anaphase transition / negative regulation of cold-induced thermogenesis / SWI/SNF complex / positive regulation of double-strand break repair / regulation of synaptic vesicle endocytosis / positive regulation of T cell differentiation / apical junction complex / structural constituent of muscle / negative regulation of calcineurin-NFAT signaling cascade / regulation of aerobic respiration / establishment or maintenance of cell polarity / cortical actin cytoskeleton / regulation of cyclin-dependent protein serine/threonine kinase activity / maintenance of blood-brain barrier / cortical cytoskeleton / positive regulation of stem cell population maintenance / NuA4 histone acetyltransferase complex / nitric-oxide synthase binding / pseudopodium / regulation of G1/S transition of mitotic cell cycle / Recycling pathway of L1 / kinesin binding / brush border / calyx of Held / negative regulation of cell differentiation / positive regulation of double-strand break repair via homologous recombination / EPH-ephrin mediated repulsion of cells / RHO GTPases Activate WASPs and WAVEs / RHO GTPases activate IQGAPs / regulation of protein localization to plasma membrane / positive regulation of myoblast differentiation / EPHB-mediated forward signaling / substantia nigra development / axonogenesis / negative regulation of protein binding / cell projection / Translocation of SLC2A4 (GLUT4) to the plasma membrane / actin filament / cell motility / RHO GTPases Activate Formins / positive regulation of cell differentiation / adherens junction / FCGR3A-mediated phagocytosis / regulation of transmembrane transporter activity / Hydrolases; Acting on acid anhydrides; Acting on acid anhydrides to facilitate cellular and subcellular movement / Signaling by high-kinase activity BRAF mutants / DNA Damage Recognition in GG-NER / MAP2K and MAPK activation / B-WICH complex positively regulates rRNA expression Similarity search - Function | ||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||
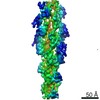
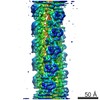
| Method | ELECTRON MICROSCOPY / helical reconstruction / cryo EM / Resolution: 15 Å | ||||||
 Authors Authors | Galkin, V.E. / Orlova, A. / Salmazo, A. / Djinovic-Carugo, K. / Egelman, E.H. | ||||||
 Citation Citation | Journal: Nat Struct Mol Biol / Year: 2010 Title: Opening of tandem calponin homology domains regulates their affinity for F-actin. Authors: Vitold E Galkin / Albina Orlova / Anita Salmazo / Kristina Djinovic-Carugo / Edward H Egelman /  Abstract: Many actin-binding proteins contain calponin homology (CH) domains, but the manner in which these domains interact with F-actin has been controversial. Crystal structures have shown the tandem CH ...Many actin-binding proteins contain calponin homology (CH) domains, but the manner in which these domains interact with F-actin has been controversial. Crystal structures have shown the tandem CH domains of alpha-actinin to be in a compact, closed conformation, but the interpretations of complexes of such tandem CH domains with F-actin have been ambiguous. We show that the tandem CH domains of alpha-actinin bind F-actin in an open conformation, explaining mutations that cause human diseases and suggesting that the opening of these domains may be one of the main regulatory mechanisms for proteins with tandem CH domains. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 3lue.cif.gz | 824.1 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb3lue.ent.gz | 640.8 KB | Display | PDB format |
| PDBx/mmJSON format | 3lue.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Summary document | 3lue_validation.pdf.gz | 861.1 KB | Display | wwPDB validaton report |
|---|---|---|---|---|
| Full document | 3lue_full_validation.pdf.gz | 1.3 MB | Display | |
| Data in XML | 3lue_validation.xml.gz | 184.5 KB | Display | |
| Data in CIF | 3lue_validation.cif.gz | 254.2 KB | Display | |
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/lu/3lueftp://data.pdbj.org/pub/pdb/validation_reports/lu/3lue | HTTPS FTP |
-Related structure data
| Related structure data |  5170MC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |



-Links
 PDBj
PDBj
















- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
| Details | Authors state that the model is from a continuous helix where the rotation per subunit is -167.2 degrees and the rise per subunit is 26.6 Angstroms. |
-Components
| #1: Protein | Mass: 41651.465 Da / Num. of mol.: 10 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: ACTB / Production host:  #2: Protein | Mass: 12471.712 Da / Num. of mol.: 10 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: ACTN3 / Production host: |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: FILAMENT / 3D reconstruction method: helical reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Tecnai F20 / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TECNAI F20 |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 200 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 200 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 50000 X / Camera length: 0 mm |
| Specimen holder | Specimen holder model: GATAN LIQUID NITROGEN / Specimen holder type: gatan 626 / Tilt angle max: 0 ° / Tilt angle min: 0 ° |
| Image recording | Film or detector model: KODAK SO-163 FILM |
| Image scans | Sampling size: 12.7 µm / Scanner model: NIKON COOLSCAN |
- Processing
Processing
| EM software |
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Details: Weiner filter | ||||||||||||
| Helical symmerty | Angular rotation/subunit: 166.8 ° / Axial rise/subunit: 27.7 Å / Axial symmetry: C1 | ||||||||||||
| 3D reconstruction | Method: back projection / Resolution: 15 Å / Resolution method: FSC 0.5 CUT-OFF / Nominal pixel size: 2.38 Å / Actual pixel size: 2.38 Å Details: AUTHORS STATE THAT THE STRANGE C-N BONDS WERE THE RESULT OF BREAKING THE CHAINS AT THESE POINTS TO DO RIGID BODY FITTING OF THE SUBDOMAINS. Symmetry type: HELICAL | ||||||||||||
| Atomic model building | Protocol: OTHER / Space: REAL Details: METHOD--both manually and with Chimera DETAILS--AUTHORS STATE THAT THE STRANGE C-N BONDS WERE THE RESULT OF BREAKING THE CHAINS AT THESE POINTS TO DO RIGID BODY FITTING OF THE SUBDOMAINS. | ||||||||||||
| Refinement step | Cycle: LAST
|
