 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 8jjs | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Human K-Ras G12D (GDP-bound) in complex with cyclic peptide inhibitor AP10343 | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード |  SIGNALING PROTEIN/INHIBITOR / CYCLIC PEPTIDE (環状ペプチド) / ONCOLOGY (腫瘍学) / SIGNALING PROTEIN-INHIBITOR COMPLEX SIGNALING PROTEIN/INHIBITOR / CYCLIC PEPTIDE (環状ペプチド) / ONCOLOGY (腫瘍学) / SIGNALING PROTEIN-INHIBITOR COMPLEX | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報forebrain astrocyte development / regulation of synaptic transmission, GABAergic / negative regulation of epithelial cell differentiation / epithelial tube branching involved in lung morphogenesis / type I pneumocyte differentiation / Rac protein signal transduction / skeletal muscle cell differentiation / positive regulation of Rac protein signal transduction / Signaling by RAS GAP mutants / Signaling by RAS GTPase mutants ...forebrain astrocyte development / regulation of synaptic transmission, GABAergic / negative regulation of epithelial cell differentiation / epithelial tube branching involved in lung morphogenesis / type I pneumocyte differentiation / Rac protein signal transduction / skeletal muscle cell differentiation / positive regulation of Rac protein signal transduction / Signaling by RAS GAP mutants / Signaling by RAS GTPase mutants / Activation of RAS in B cells / RAS signaling downstream of NF1 loss-of-function variants / RUNX3 regulates p14-ARF / SOS-mediated signalling / Activated NTRK3 signals through RAS / Activated NTRK2 signals through RAS / SHC1 events in ERBB4 signaling / Signalling to RAS / glial cell proliferation / SHC-related events triggered by IGF1R / Activated NTRK2 signals through FRS2 and FRS3 / Estrogen-stimulated signaling through PRKCZ / SHC-mediated cascade:FGFR3 / MET activates RAS signaling / PTK6 Regulates RHO GTPases, RAS GTPase and MAP kinases / Signaling by PDGFRA transmembrane, juxtamembrane and kinase domain mutants / Signaling by PDGFRA extracellular domain mutants / SHC-mediated cascade:FGFR2 / SHC-mediated cascade:FGFR4 / Signaling by FGFR4 in disease / SHC-mediated cascade:FGFR1 / Erythropoietin activates RAS / protein-membrane adaptor activity / homeostasis of number of cells within a tissue / positive regulation of glial cell proliferation / FRS-mediated FGFR3 signaling / Signaling by CSF3 (G-CSF) / Signaling by FLT3 ITD and TKD mutants / FRS-mediated FGFR2 signaling / FRS-mediated FGFR4 signaling / Signaling by FGFR3 in disease / FRS-mediated FGFR1 signaling / p38MAPK events / Tie2 Signaling / Signaling by FGFR2 in disease / striated muscle cell differentiation / GRB2 events in EGFR signaling / SHC1 events in EGFR signaling / EGFR Transactivation by Gastrin / Signaling by FLT3 fusion proteins / FLT3 Signaling / Ras activation upon Ca2+ influx through NMDA receptor / GRB2 events in ERBB2 signaling / Signaling by FGFR1 in disease / NCAM signaling for neurite out-growth / CD209 (DC-SIGN) signaling / SHC1 events in ERBB2 signaling / Downstream signal transduction / Constitutive Signaling by Overexpressed ERBB2 / Insulin receptor signalling cascade / 低分子量GTPアーゼ / G protein activity / Signaling by phosphorylated juxtamembrane, extracellular and kinase domain KIT mutants / VEGFR2 mediated cell proliferation / FCERI mediated MAPK activation / regulation of long-term neuronal synaptic plasticity / Signaling by ERBB2 TMD/JMD mutants / RAF activation / Signaling by high-kinase activity BRAF mutants / Constitutive Signaling by EGFRvIII / visual learning / MAP2K and MAPK activation / Signaling by ERBB2 ECD mutants / Signaling by ERBB2 KD Mutants / Signaling by SCF-KIT / cytoplasmic side of plasma membrane / Regulation of RAS by GAPs / Negative regulation of MAPK pathway / RAS processing / GDP binding / Signaling by RAF1 mutants / Signaling by moderate kinase activity BRAF mutants / Paradoxical activation of RAF signaling by kinase inactive BRAF / Signaling downstream of RAS mutants / Signaling by CSF1 (M-CSF) in myeloid cells / 分裂促進因子活性化タンパク質キナーゼ / Signaling by BRAF and RAF1 fusions / Constitutive Signaling by Ligand-Responsive EGFR Cancer Variants / DAP12 signaling / Ca2+ pathway / 遺伝子発現 / actin cytoskeleton organization / RAF/MAP kinase cascade / neuron apoptotic process / Ras protein signal transduction / negative regulation of neuron apoptotic process / mitochondrial outer membrane / positive regulation of protein phosphorylation / ゴルジ体 / focal adhesion類似検索 - 分子機能 | ||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト)synthetic construct (人工物) | ||||||
| 手法 | X線回折 / シンクロトロン / 分子置換 / 解像度: 1.534 Å | ||||||
 データ登録者 データ登録者 | Irie, M. / Fukami, T.A. / Tanada, M. / Ohta, A. / Torizawa, T. | ||||||
| 資金援助 | 1件
| ||||||
 引用 引用 | ジャーナル: J.Am.Chem.Soc. / 年: 2023 タイトル: Development of Orally Bioavailable Peptides Targeting an Intracellular Protein: From a Hit to a Clinical KRAS Inhibitor. 著者: Tanada, M. / Tamiya, M. / Matsuo, A. / Chiyoda, A. / Takano, K. / Ito, T. / Irie, M. / Kotake, T. / Takeyama, R. / Kawada, H. / Hayashi, R. / Ishikawa, S. / Nomura, K. / Furuichi, N. / ...著者: Tanada, M. / Tamiya, M. / Matsuo, A. / Chiyoda, A. / Takano, K. / Ito, T. / Irie, M. / Kotake, T. / Takeyama, R. / Kawada, H. / Hayashi, R. / Ishikawa, S. / Nomura, K. / Furuichi, N. / Morita, Y. / Kage, M. / Hashimoto, S. / Nii, K. / Sase, H. / Ohara, K. / Ohta, A. / Kuramoto, S. / Nishimura, Y. / Iikura, H. / Shiraishi, T. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8jjs.cif.gz | 62.4 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8jjs.ent.gz | 39.2 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 8jjs.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/jj/8jjsftp://data.pdbj.org/pub/pdb/validation_reports/jj/8jjs | HTTPS FTP |
|---|
-関連構造データ
| 類似構造データ |
|---|
-リンク
 PDBj
PDBj




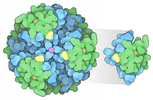









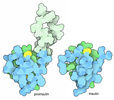


- 集合体
集合体
| 登録構造単位 | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||
| 単位格子 |
|
-要素
-タンパク質 / タンパク質・ペプチド , 2種, 2分子 AI
| #1: タンパク質 | 分子量: 20150.582 Da / 分子数: 1 / 変異: G12D / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 発現宿主:  Escherichia coli (大腸菌) / 株 (発現宿主): Tuner(DE3)pLacI / 参照: UniProt: P01116 Escherichia coli (大腸菌) / 株 (発現宿主): Tuner(DE3)pLacI / 参照: UniProt: P01116 |
|---|---|
| #2: タンパク質・ペプチド | 分子量: 1424.091 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
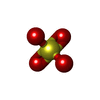
-非ポリマー , 4種, 127分子 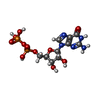


| #3: 化合物 | ChemComp-GDP / グアノシン二リン酸 タイプ: RNA linking / 分子量: 443.201 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H15N5O11P2 / コメント: GDP, エネルギー貯蔵分子*YM タイプ: RNA linking / 分子量: 443.201 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H15N5O11P2 / コメント: GDP, エネルギー貯蔵分子*YM | ||
|---|---|---|---|
| #4: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION 分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION | ||
| #5: 化合物 | 硫酸塩 分子量: 96.063 Da / 分子数: 3 / 由来タイプ: 合成 / 式: SO4 / タイプ: SUBJECT OF INVESTIGATION 分子量: 96.063 Da / 分子数: 3 / 由来タイプ: 合成 / 式: SO4 / タイプ: SUBJECT OF INVESTIGATION#6: 水 | ChemComp-HOH / | 水分子量: 18.015 Da / 分子数: 122 / 由来タイプ: 天然 / 式: H2O |
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|
-実験情報
-実験
| 実験 | 手法: X線回折 / 使用した結晶の数: 1 |
|---|
- 試料調製
試料調製
| 結晶 | マシュー密度: 2.57 Å3/Da / 溶媒含有率: 52.14 % |
|---|---|
| 結晶化 | 温度: 293 K / 手法: 蒸気拡散法, シッティングドロップ法 詳細: 2.0 M Ammonium sulfate, 25 %(v/v) Ethylene glycol as cryoprotectant |
-データ収集
| 回折 | 平均測定温度: 95 K / Serial crystal experiment: N | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 放射光源 | 由来: シンクロトロン / サイト: Photon Factory  / ビームライン: BL-1A / 波長: 1.045 Å / ビームライン: BL-1A / 波長: 1.045 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 検出器 | タイプ: DECTRIS EIGER X 4M / 検出器: PIXEL / 日付: 2021年12月13日 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 放射 | プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 放射波長 | 波長: 1.045 Å / 相対比: 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 反射 | 解像度: 1.534→57.674 Å / Num. obs: 21593 / % possible obs: 95 % / 冗長度: 20.49 % 詳細: Some remarks regarding the mmCIF items written, the PDB Exchange Dictionary (PDBx/mmCIF) Version 5.0 supporting the data files in the current PDB archive (dictionary version 5.325, last ...詳細: Some remarks regarding the mmCIF items written, the PDB Exchange Dictionary (PDBx/mmCIF) Version 5.0 supporting the data files in the current PDB archive (dictionary version 5.325, last updated 2020-04-13: http://mmcif.wwpdb.org/dictionaries/mmcif_pdbx_v50.dic/Index/) and the actual quantities provided by MRFANA (https://github.com/githubgphl/MRFANA) from the autoPROC package (https://www.globalphasing.com/autoproc/). In general, the mmCIF categories here should provide items that are currently used in the PDB archive. If there are alternatives, the one recommended by the PDB developers has been selected. The distinction between *_all and *_obs quantities is not always clear: often only one version is actively used within the PDB archive (or is the one recommended by PDB developers). The intention of distinguishing between classes of reflections before and after some kind of observation criterion was applied, can in principle be useful - but such criteria change in various ways throughout the data processing steps (rejection of overloaded or too partial reflections, outlier/misfit rejections during scaling etc) and there is no retrospect computation of data scaling/merging statistics for the reflections used in the final refinement (where another observation criterion might have been applied). Typical data processing will usually only provide one version of statistics at various stages and these are given in the recommended item here, irrespective of the "_all" and "_obs" connotation, see e.g. the use of _reflns.pdbx_Rmerge_I_obs, _reflns.pdbx_Rrim_I_all and _reflns.pdbx_Rpim_I_all. Please note that all statistics related to "merged intensities" (or "merging") are based on inverse-variance weighting of the individual measurements making up a symmetry-unique reflection. This is standard for several decades now, even if some of the dictionary definitions seem to suggest that a simple "mean" or "average" intensity is being used instead. R-values are always given for all symmetry-equivalent reflections following Friedel's law, i.e. Bijvoet pairs are not treated separately (since we want to describe the overall mean intensity and not the mean I(+) and I(-) here). The Rrim metric is identical to the Rmeas R-value and only differs in name. _reflns.pdbx_number_measured_all is the number of measured intensities just before the final merging step (at which point no additional rejection takes place). _reflns.number_obs is the number of symmetry-unique observations, i.e. the result of merging those measurements via inverse-variance weighting. _reflns.pdbx_netI_over_sigmaI is based on the merged intensities (_reflns.number_obs) as expected. _reflns.pdbx_redundancy is synonymous with "multiplicity". The per-shell item _reflns_shell.number_measured_all corresponds to the overall value _reflns.pdbx_number_measured_all. The per-shell item _reflns_shell.number_unique_all corresponds to the overall value _reflns.number_obs. The per-shell item _reflns_shell.percent_possible_all corresponds to the overall value _reflns.percent_possible_obs. The per-shell item _reflns_shell.meanI_over_sigI_obs corresponds to the overall value given as _reflns.pdbx_netI_over_sigmaI. But be aware of the incorrect definition of the former in the current dictionary! CC1/2: 0.999 / CC1/2 anomalous: -0.329 / Rmerge(I) obs: 0.104 / Rpim(I) all: 0.0232 / Rrim(I) all: 0.1065 / AbsDiff over sigma anomalous: 0.743 / Baniso tensor eigenvalue 1: 30.8 Å2 / Baniso tensor eigenvalue 2: 30.8 Å2 / Baniso tensor eigenvalue 3: 17.8 Å2 / Baniso tensor eigenvector 1 ortho1: 1 / Baniso tensor eigenvector 1 ortho2: 0 / Baniso tensor eigenvector 1 ortho3: 0 / Baniso tensor eigenvector 2 ortho1: 0 / Baniso tensor eigenvector 2 ortho2: 1 / Baniso tensor eigenvector 2 ortho3: 0 / Baniso tensor eigenvector 3 ortho1: 0 / Baniso tensor eigenvector 3 ortho2: 0 / Baniso tensor eigenvector 3 ortho3: 1 / Aniso diffraction limit 1: 1.856 Å / Aniso diffraction limit 2: 1.856 Å / Aniso diffraction limit 3: 1.499 Å / Aniso diffraction limit axis 1 ortho1: 1 / Aniso diffraction limit axis 1 ortho2: 0 / Aniso diffraction limit axis 1 ortho3: 0 / Aniso diffraction limit axis 2 ortho1: 0 / Aniso diffraction limit axis 2 ortho2: 1 / Aniso diffraction limit axis 2 ortho3: 0 / Aniso diffraction limit axis 3 ortho1: 0 / Aniso diffraction limit axis 3 ortho2: 0 / Aniso diffraction limit axis 3 ortho3: 1 / Net I/σ(I): 19.1 / Num. measured all: 442425 / Orthogonalization convention: pdb / % possible anomalous: 94.4 / % possible ellipsoidal: 95 / % possible ellipsoidal anomalous: 94.4 / % possible spherical: 66.2 / % possible spherical anomalous: 65.8 / Redundancy anomalous: 10.42 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 反射 シェル |
|
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 精密化 | 構造決定の手法: 分子置換 / 解像度: 1.534→57.67 Å / Cor.coef. Fo:Fc: 0.941 / Cor.coef. Fo:Fc free: 0.932 / SU R Cruickshank DPI: 0.126 / 交差検証法: THROUGHOUT / σ(F): 0 / SU R Blow DPI: 0.128 / SU Rfree Blow DPI: 0.116 / SU Rfree Cruickshank DPI: 0.116
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso max: 77.55 Å2 / Biso mean: 27.04 Å2 / Biso min: 10.44 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine analyze | Luzzati coordinate error obs: 0.26 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: final / 解像度: 1.534→57.67 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS精密化 シェル | 解像度: 1.534→1.62 Å / Rfactor Rfree error: 0
|
