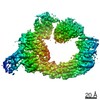
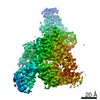
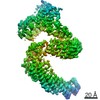
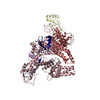
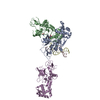
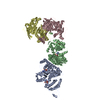
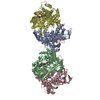
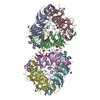
登録情報 データベース : PDB / ID : 7b5sタイトル Ubiquitin ligation to F-box protein substrates by SCF-RBR E3-E3 super-assembly: CUL1-RBX1-ARIH1 Ariadne. Transition State 1 Cullin-1 E3 ubiquitin-protein ligase ARIH1 E3 ubiquitin-protein ligase RBX1 キーワード / / / / / / / / / / / 機能・相同性 分子機能 ドメイン・相同性 構成要素
/ / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / / 生物種 Homo sapiens (ヒト)手法 / / / 解像度 : 3.6 Å データ登録者 Horn-Ghetko, D. / Prabu, J.R. / Schulman, B.A. 資金援助 組織 認可番号 国 European Research Council (ERC) H2020 789016-NEDD8Activate German Research Foundation (DFG) SCHU 3196/1-1
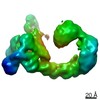
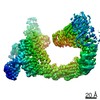
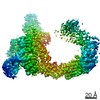
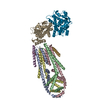
ジャーナル : Nature / 年 : 2021タイトル : Ubiquitin ligation to F-box protein targets by SCF-RBR E3-E3 super-assembly.著者 : Daniel Horn-Ghetko / David T Krist / J Rajan Prabu / Kheewoong Baek / Monique P C Mulder / Maren Klügel / Daniel C Scott / Huib Ovaa / Gary Kleiger / Brenda A Schulman / 要旨 : E3 ligases are typically classified by hallmark domains such as RING and RBR, which are thought to specify unique catalytic mechanisms of ubiquitin transfer to recruited substrates. However, rather ... E3 ligases are typically classified by hallmark domains such as RING and RBR, which are thought to specify unique catalytic mechanisms of ubiquitin transfer to recruited substrates. However, rather than functioning individually, many neddylated cullin-RING E3 ligases (CRLs) and RBR-type E3 ligases in the ARIH family-which together account for nearly half of all ubiquitin ligases in humans-form E3-E3 super-assemblies. Here, by studying CRLs in the SKP1-CUL1-F-box (SCF) family, we show how neddylated SCF ligases and ARIH1 (an RBR-type E3 ligase) co-evolved to ubiquitylate diverse substrates presented on various F-box proteins. We developed activity-based chemical probes that enabled cryo-electron microscopy visualization of steps in E3-E3 ubiquitylation, initiating with ubiquitin linked to the E2 enzyme UBE2L3, then transferred to the catalytic cysteine of ARIH1, and culminating in ubiquitin linkage to a substrate bound to the SCF E3 ligase. The E3-E3 mechanism places the ubiquitin-linked active site of ARIH1 adjacent to substrates bound to F-box proteins (for example, substrates with folded structures or limited length) that are incompatible with previously described conventional RING E3-only mechanisms. The versatile E3-E3 super-assembly may therefore underlie widespread ubiquitylation. 履歴 登録 2020年12月7日 登録サイト / 処理サイト 改定 1.0 2021年2月10日 Provider / タイプ 改定 1.1 2021年2月17日 Group / カテゴリ / citation_authorItem / _citation.title / _citation_author.name改定 1.2 2021年3月10日 Group / カテゴリ Item / _citation.page_first / _citation.page_last改定 1.3 2024年5月1日 Group / Database references / カテゴリ / chem_comp_bond / database_2Item / _database_2.pdbx_database_accession
すべて表示 表示を減らす
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード LIGASE (リガーゼ) /
LIGASE (リガーゼ) /  機能・相同性情報
機能・相同性情報
 データ登録者
データ登録者 ドイツ, 2件
ドイツ, 2件  引用
引用

 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード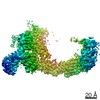











 PDBj
PDBj








 集合体
集合体



 分子量: 65.409 Da / 分子数: 5 / 由来タイプ: 合成 / 式: Zn
分子量: 65.409 Da / 分子数: 5 / 由来タイプ: 合成 / 式: Zn 試料調製
試料調製 電子顕微鏡撮影
電子顕微鏡撮影
 解析
解析