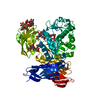
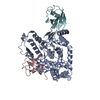
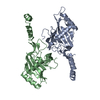
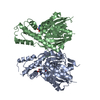
- PDB-6i2v: Pilotin from Vibrio vulnificus type 2 secretion system, EpsS. -
+
Open data
ID or keywords:
Loading...
-
Basic information
Entry
Database: PDB / ID: 6i2v
Title
Pilotin from Vibrio vulnificus type 2 secretion system, EpsS.
Components
Uncharacterized protein
Keywords
PROTEIN BINDING / Pilotin / T2SS / V.vulnificus / Outer-membrane
Function / homology
GMP Synthetase; Chain A, domain 3 - #250 / Type II secretion system (T2SS) pilotin, S protein / Type II secretion system (T2SS) pilotin, S protein / GMP Synthetase; Chain A, domain 3 / 2-Layer Sandwich / Alpha Beta / SULFITE ION / Uncharacterized protein
Journal: PLoS Pathog / Year: 2019 Title: Structure and assembly of pilotin-dependent and -independent secretins of the type II secretion system. Authors: S Peter Howard / Leandro F Estrozi / Quentin Bertrand / Carlos Contreras-Martel / Timothy Strozen / Viviana Job / Alexandre Martins / Daphna Fenel / Guy Schoehn / Andréa Dessen / Abstract: The type II secretion system (T2SS) is a cell envelope-spanning macromolecular complex that is prevalent in Gram-negative bacterial species. It serves as the predominant virulence mechanism of many ...The type II secretion system (T2SS) is a cell envelope-spanning macromolecular complex that is prevalent in Gram-negative bacterial species. It serves as the predominant virulence mechanism of many bacteria including those of the emerging human pathogens Vibrio vulnificus and Aeromonas hydrophila. The system is composed of a core set of highly conserved proteins that assemble an inner membrane platform, a periplasmic pseudopilus and an outer membrane complex termed the secretin. Localization and assembly of secretins in the outer membrane requires recognition of secretin monomers by two different partner systems: an inner membrane accessory complex or a highly sequence-diverse outer membrane lipoprotein, termed the pilotin. In this study, we addressed the question of differential secretin assembly mechanisms by using cryo-electron microscopy to determine the structures of the secretins from A. hydrophila (pilotin-independent ExeD) and V. vulnificus (pilotin-dependent EpsD). These structures, at approximately 3.5 Å resolution, reveal pentadecameric stoichiometries and C-terminal regions that carry a signature motif in the case of a pilotin-dependent assembly mechanism. We solved the crystal structure of the V. vulnificus EpsS pilotin and confirmed the importance of the signature motif for pilotin-dependent secretin assembly by performing modelling with the C-terminus of EpsD. We also show that secretin assembly is essential for membrane integrity and toxin secretion in V. vulnificus and establish that EpsD requires the coordinated activity of both the accessory complex EpsAB and the pilotin EpsS for full assembly and T2SS function. In contrast, mutation of the region of the S-domain that is normally the site of pilotin interactions has little effect on assembly or function of the ExeD secretin. Since secretins are essential outer membrane channels present in a variety of secretion systems, these results provide a structural and functional basis for understanding the key assembly steps for different members of this vast pore-forming family of proteins.
Resolution: 1.75→45.39 Å / Cor.coef. Fo:Fc: 0.964 / Cor.coef. Fo:Fc free: 0.939 / SU B: 5.128 / SU ML: 0.081 / Cross valid method: THROUGHOUT / ESU R: 0.12 / ESU R Free: 0.123 / Details: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
Rfactor
Num. reflection
% reflection
Selection details
Rfree
0.22787
1822
13 %
RANDOM
Rwork
0.17904
-
-
-
obs
0.18546
12215
99.38 %
-
Solvent computation
Ion probe radii: 0.8 Å / Shrinkage radii: 0.8 Å / VDW probe radii: 1.2 Å
 Movie
Movie Controller
Controller


 Open data
Open data
 Basic information
Basic information Components
Components Keywords
Keywords PROTEIN BINDING / Pilotin /
PROTEIN BINDING / Pilotin /  Function and homology information
Function and homology information
 Authors
Authors Citation
Citation


 Structure visualization
Structure visualization Downloads & links
Downloads & links Other downloads
Other downloads








 PDBj
PDBj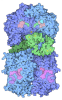
 Assembly
Assembly

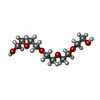


 Mass: 238.278 Da / Num. of mol.: 5 / Source method: obtained synthetically / Formula: C10H22O6 / Comment: precipitant*YM
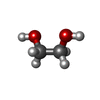
Mass: 238.278 Da / Num. of mol.: 5 / Source method: obtained synthetically / Formula: C10H22O6 / Comment: precipitant*YM Mass: 62.068 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C2H6O2
Mass: 62.068 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C2H6O2 Mass: 80.063 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: SO3
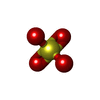
Mass: 80.063 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: SO3 Mass: 96.063 Da / Num. of mol.: 5 / Source method: obtained synthetically / Formula: SO4
Mass: 96.063 Da / Num. of mol.: 5 / Source method: obtained synthetically / Formula: SO4 Sample preparation
Sample preparation Processing
Processing