 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
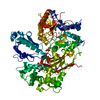
Yorodumi- PDB-5fwx: Crystal structure of the AMPA receptor GluA2/A4 N-terminal domain... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 5fwx | ||||||
|---|---|---|---|---|---|---|---|
| Title | Crystal structure of the AMPA receptor GluA2/A4 N-terminal domain heterodimer | ||||||
 Components Components |
| ||||||
 Keywords Keywords |  TRANSPORT PROTEIN TRANSPORT PROTEIN | ||||||
| Function / homology |  Function and homology information Function and homology informationTrafficking of AMPA receptors / kainate selective glutamate receptor complex / regulation of synapse structure or activity / Synaptic adhesion-like molecules / spine synapse / dendritic spine neck / dendritic spine head / Activation of AMPA receptors / response to lithium ion / perisynaptic space ...Trafficking of AMPA receptors / kainate selective glutamate receptor complex / regulation of synapse structure or activity / Synaptic adhesion-like molecules / spine synapse / dendritic spine neck / dendritic spine head / Activation of AMPA receptors / response to lithium ion / perisynaptic space / cellular response to glycine / AMPA glutamate receptor activity / negative regulation of smooth muscle cell apoptotic process / Trafficking of GluR2-containing AMPA receptors / immunoglobulin binding / AMPA glutamate receptor complex / kainate selective glutamate receptor activity / ionotropic glutamate receptor complex / extracellularly glutamate-gated ion channel activity / asymmetric synapse / regulation of receptor recycling / Unblocking of NMDA receptors, glutamate binding and activation / glutamate receptor binding / positive regulation of synaptic transmission / presynaptic active zone membrane / glutamate-gated receptor activity / response to fungicide / regulation of synaptic transmission, glutamatergic / cellular response to brain-derived neurotrophic factor stimulus / somatodendritic compartment / dendrite membrane / ligand-gated monoatomic ion channel activity involved in regulation of presynaptic membrane potential / ionotropic glutamate receptor binding / ionotropic glutamate receptor signaling pathway / cytoskeletal protein binding / dendrite cytoplasm / positive regulation of synaptic transmission, glutamatergic / SNARE binding / dendritic shaft / transmitter-gated monoatomic ion channel activity involved in regulation of postsynaptic membrane potential / synaptic membrane / synaptic transmission, glutamatergic / PDZ domain binding / postsynaptic density membrane / protein tetramerization / Schaffer collateral - CA1 synapse / modulation of chemical synaptic transmission / establishment of protein localization / terminal bouton / receptor internalization / cerebral cortex development / synaptic vesicle membrane / synaptic vesicle / presynapse / presynaptic membrane / signaling receptor activity / amyloid-beta binding / growth cone / perikaryon / scaffold protein binding / chemical synaptic transmission / postsynaptic membrane / dendritic spine / postsynaptic density / neuron projection / axon / neuronal cell body / synapse / glutamatergic synapse / dendrite / protein-containing complex binding / endoplasmic reticulum membrane / protein kinase binding / cell surface / endoplasmic reticulum / protein-containing complex / membrane / identical protein binding / plasma membraneSimilarity search - Function | ||||||
| Biological species |  RATTUS NORVEGICUS (Norway rat) RATTUS NORVEGICUS (Norway rat) | ||||||
| Method | X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.5 Å | ||||||
 Authors Authors | Garcia-Nafria, J. / Herguedas, B. / Greger, I.H. | ||||||
 Citation Citation | Journal: Science / Year: 2016 Title: Structure and organization of heteromeric AMPA-type glutamate receptors. Authors: Beatriz Herguedas / Javier García-Nafría / Ondrej Cais / Rafael Fernández-Leiro / James Krieger / Hinze Ho / Ingo H Greger /  Abstract: AMPA-type glutamate receptors (AMPARs), which are central mediators of rapid neurotransmission and synaptic plasticity, predominantly exist as heteromers of the subunits GluA1 to GluA4. Here we ...AMPA-type glutamate receptors (AMPARs), which are central mediators of rapid neurotransmission and synaptic plasticity, predominantly exist as heteromers of the subunits GluA1 to GluA4. Here we report the first AMPAR heteromer structures, which deviate substantially from existing GluA2 homomer structures. Crystal structures of the GluA2/3 and GluA2/4 N-terminal domains reveal a novel compact conformation with an alternating arrangement of the four subunits around a central axis. This organization is confirmed by cysteine cross-linking in full-length receptors, and it permitted us to determine the structure of an intact GluA2/3 receptor by cryogenic electron microscopy. Two models in the ligand-free state, at resolutions of 8.25 and 10.3 angstroms, exhibit substantial vertical compression and close associations between domain layers, reminiscent of N-methyl-D-aspartate receptors. Model 1 resembles a resting state and model 2 a desensitized state, thus providing snapshots of gating transitions in the nominal absence of ligand. Our data reveal organizational features of heteromeric AMPARs and provide a framework to decipher AMPAR architecture and signaling. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 5fwx.cif.gz | 563.5 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb5fwx.ent.gz | 462.1 KB | Display | PDB format |
| PDBx/mmJSON format | 5fwx.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/fw/5fwxftp://data.pdbj.org/pub/pdb/validation_reports/fw/5fwx | HTTPS FTP |
|---|
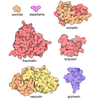
-Related structure data
| Related structure data |  8090C  8091C  5fwyC 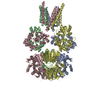 5ideC  5idfC 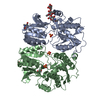 3hsyS  4gpaS S: Starting model for refinement C: citing same article ( |
|---|---|
| Similar structure data |
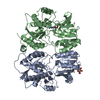
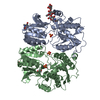


-Links
 PDBj
PDBj



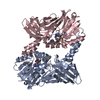
- Assembly
Assembly
| Deposited unit | 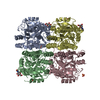
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 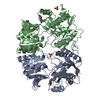
| ||||||||
| 2 | 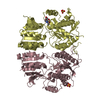
| ||||||||
| Unit cell |
|
-Components
| #1: Protein | GRIA2 / GLUR-2 / AMPA-SELECTIVE GLUTAMATE RECEPTOR 2 / GLUR-B / GLUR-K 2 / GLUTAMATE RECEPTOR IONOTROPIC / ...GLUR-2 / AMPA-SELECTIVE GLUTAMATE RECEPTOR 2 / GLUR-B / GLUR-K 2 / GLUTAMATE RECEPTOR IONOTROPIC / AMPA 2 / GLUA2 / GLUA2 AMPA RECEPTOR N-TERMINAL DOMAIN Mass: 43731.359 Da / Num. of mol.: 2 / Fragment: RESIDUES 25-400 Source method: isolated from a genetically manipulated source Source: (gene. exp.) RATTUS NORVEGICUS (Norway rat) / Cell line (production host): HEK293 GNT1 / Production host:  HOMO SAPIENS (human) / References: UniProt: P19491 HOMO SAPIENS (human) / References: UniProt: P19491#2: Protein | / GLUR-4 / GLUR4 / AMPA-SELECTIVE GLUTAMATE RECEPTOR 4 / GLUR-D / GLUTAMATE RECEPTOR IONOTROPIC / ...GLUR-4 / GLUR4 / AMPA-SELECTIVE GLUTAMATE RECEPTOR 4 / GLUR-D / GLUTAMATE RECEPTOR IONOTROPIC / AMPA 4 / GLUA4 / GLUA4 AMPA RECEPTOR N-TERMINAL DOMAINMass: 44547.430 Da / Num. of mol.: 2 / Fragment: RESIDUES 22-401 Source method: isolated from a genetically manipulated source Source: (gene. exp.) RATTUS NORVEGICUS (Norway rat) / Cell line (production host): HEK293 GNT1 / Production host: HOMO SAPIENS (human) / References: UniProt: P19493#3: Chemical | ChemComp-SO4 / Sulfate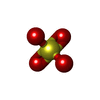  Mass: 96.063 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: SO4 Mass: 96.063 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: SO4#4: Sugar | N-Acetylglucosamine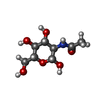  Type: D-saccharide, beta linking / Mass: 221.208 Da / Num. of mol.: 3 Type: D-saccharide, beta linking / Mass: 221.208 Da / Num. of mol.: 3Source method: isolated from a genetically manipulated source Formula: C8H15NO6 #5: Water | ChemComp-HOH / | Water Mass: 18.015 Da / Num. of mol.: 125 / Source method: isolated from a natural source / Formula: H2O Mass: 18.015 Da / Num. of mol.: 125 / Source method: isolated from a natural source / Formula: H2O |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 2.5 Å3/Da / Density % sol: 40.01 % / Description: NONE |
|---|---|
| Crystal grow | Details: 17% PEG 3350 WITH 0.1-0.35 M AMMONIUM SULPHATE |
-Data collection
| Diffraction | Mean temperature: 100 K | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: Diamond / Beamline: I04-1 / Wavelength: 0.92 | |||||||||||||||
| Detector | Type: DECTRIS PILATUS / Detector: PIXEL / Date: Dec 9, 2012 | |||||||||||||||
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray | |||||||||||||||
| Radiation wavelength | Wavelength: 0.92 Å / Relative weight: 1 | |||||||||||||||
| Reflection twin |
| |||||||||||||||
| Reflection | Resolution: 2.5→39.68 Å / Num. obs: 45771 / % possible obs: 96.3 % / Observed criterion σ(I): 2 / Redundancy: 3.2 % / Rmerge(I) obs: 0.06 / Net I/σ(I): 12.2 | |||||||||||||||
| Reflection shell | Resolution: 2.5→2.64 Å / Redundancy: 3.3 % / Rmerge(I) obs: 0.35 / Mean I/σ(I) obs: 3 / % possible all: 96 |
- Processing
Processing
| Software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: PDB ENTRIES 4GPA AND 3HSY Resolution: 2.5→39.16 Å / Cor.coef. Fo:Fc: 0.962 / Cor.coef. Fo:Fc free: 0.936 / SU B: 15.791 / SU ML: 0.187 / Cross valid method: THROUGHOUT / ESU R: 0.499 / ESU R Free: 0.065 / Stereochemistry target values: MAXIMUM LIKELIHOOD Details: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS. HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS U VALUES WITH TLS ADDED
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Ion probe radii: 0.8 Å / Shrinkage radii: 0.8 Å / VDW probe radii: 1.2 Å / Solvent model: MASK | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 58.23 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 2.5→39.16 Å
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
|