regulation of protein deubiquitination / negative regulation of cytoplasmic pattern recognition receptor signaling pathway / nucleotide-binding domain, leucine rich repeat containing receptor signaling pathway / ubiquitin-like protein transferase activity / negative regulation of defense response to virus / protein K29-linked ubiquitination / T cell anergy / positive regulation of T cell anergy / CXCR chemokine receptor binding / regulation of necroptotic process ...regulation of protein deubiquitination / negative regulation of cytoplasmic pattern recognition receptor signaling pathway / nucleotide-binding domain, leucine rich repeat containing receptor signaling pathway / ubiquitin-like protein transferase activity / negative regulation of defense response to virus / protein K29-linked ubiquitination / T cell anergy / positive regulation of T cell anergy / CXCR chemokine receptor binding / regulation of necroptotic process / CD4-positive, alpha-beta T cell proliferation / negative regulation of CD4-positive, alpha-beta T cell proliferation / negative regulation of JNK cascade / HECT-type E3 ubiquitin transferase / arrestin family protein binding / regulation of hematopoietic stem cell differentiation / negative regulation of type I interferon production / positive regulation of receptor catabolic process / negative regulation of NF-kappaB transcription factor activity / ubiquitin-like protein ligase binding / protein monoubiquitination / ligase activity / protein K63-linked ubiquitination / protein autoubiquitination / protein K48-linked ubiquitination / ribonucleoprotein complex binding / Maturation of protein E / Maturation of protein E / ER Quality Control Compartment (ERQC) / Myoclonic epilepsy of Lafora / FLT3 signaling by CBL mutants / Prevention of phagosomal-lysosomal fusion / IRAK2 mediated activation of TAK1 complex / Alpha-protein kinase 1 signaling pathway / グリコーゲン合成 / IRAK1 recruits IKK complex / IRAK1 recruits IKK complex upon TLR7/8 or 9 stimulation / Membrane binding and targetting of GAG proteins / Endosomal Sorting Complex Required For Transport (ESCRT) / IRAK2 mediated activation of TAK1 complex upon TLR7/8 or 9 stimulation / PTK6 Regulates RTKs and Their Effectors AKT1 and DOK1 / Negative regulation of FLT3 / Constitutive Signaling by NOTCH1 HD Domain Mutants / Regulation of FZD by ubiquitination / TICAM1,TRAF6-dependent induction of TAK1 complex / NOTCH2 Activation and Transmission of Signal to the Nucleus / TICAM1-dependent activation of IRF3/IRF7 / APC/C:Cdc20 mediated degradation of Cyclin B / p75NTR recruits signalling complexes / Downregulation of ERBB4 signaling / TRAF6 mediated IRF7 activation in TLR7/8 or 9 signaling / APC-Cdc20 mediated degradation of Nek2A / PINK1-PRKN Mediated Mitophagy / TRAF6-mediated induction of TAK1 complex within TLR4 complex / Pexophagy / Regulation of innate immune responses to cytosolic DNA / VLDLR internalisation and degradation / InlA-mediated entry of Listeria monocytogenes into host cells / Downregulation of ERBB2:ERBB3 signaling / NF-kB is activated and signals survival / NRIF signals cell death from the nucleus / Regulation of PTEN localization / Activated NOTCH1 Transmits Signal to the Nucleus / Regulation of BACH1 activity / Translesion synthesis by REV1 / Synthesis of active ubiquitin: roles of E1 and E2 enzymes / Translesion synthesis by POLK / MAP3K8 (TPL2)-dependent MAPK1/3 activation / TICAM1, RIP1-mediated IKK complex recruitment / Downregulation of TGF-beta receptor signaling / Activation of IRF3, IRF7 mediated by TBK1, IKKε (IKBKE) / Translesion synthesis by POLI / Gap-filling DNA repair synthesis and ligation in GG-NER / Josephin domain DUBs / Regulation of activated PAK-2p34 by proteasome mediated degradation / InlB-mediated entry of Listeria monocytogenes into host cell / IKK complex recruitment mediated by RIP1 / JNK (c-Jun kinases) phosphorylation and activation mediated by activated human TAK1 / TGF-beta receptor signaling in EMT (epithelial to mesenchymal transition) / N-glycan trimming in the ER and Calnexin/Calreticulin cycle / Autodegradation of Cdh1 by Cdh1:APC/C / TNFR1-induced NF-kappa-B signaling pathway / APC/C:Cdc20 mediated degradation of Securin / Asymmetric localization of PCP proteins / SCF-beta-TrCP mediated degradation of Emi1 / TCF dependent signaling in response to WNT / NIK-->noncanonical NF-kB signaling / Regulation of NF-kappa B signaling / Ubiquitin-dependent degradation of Cyclin D / AUF1 (hnRNP D0) binds and destabilizes mRNA / TNFR2 non-canonical NF-kB pathway / activated TAK1 mediates p38 MAPK activation / Assembly of the pre-replicative complex / Vpu mediated degradation of CD4 / NOTCH3 Activation and Transmission of Signal to the Nucleus / Negative regulators of DDX58/IFIH1 signaling / Deactivation of the beta-catenin transactivating complex / Degradation of DVL / Ubiquitin Mediated Degradation of Phosphorylated Cdc25A / Regulation of signaling by CBL 類似検索 - 分子機能
E3 ubiquitin-protein ligase, SMURF1 type / HECT domain / HECT, E3 ligase catalytic domain / HECT-domain (ubiquitin-transferase) / HECT domain profile. / Domain Homologous to E6-AP Carboxyl Terminus with / C2ドメイン / Protein kinase C conserved region 2 (CalB) / WWドメイン / WW/rsp5/WWP domain signature. ...E3 ubiquitin-protein ligase, SMURF1 type / HECT domain / HECT, E3 ligase catalytic domain / HECT-domain (ubiquitin-transferase) / HECT domain profile. / Domain Homologous to E6-AP Carboxyl Terminus with / C2ドメイン / Protein kinase C conserved region 2 (CalB) / WWドメイン / WW/rsp5/WWP domain signature. / C2ドメイン / C2 domain profile. / WW domain superfamily / WW/rsp5/WWP domain profile. / Domain with 2 conserved Trp (W) residues / WWドメイン / C2 domain superfamily / Ubiquitin domain signature. / Ubiquitin conserved site / Ubiquitin domain / Ubiquitin family / Ubiquitin homologues / Ubiquitin domain profile. / ユビキチン様タンパク質 / Ubiquitin-like domain superfamily 類似検索 - ドメイン・相同性
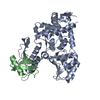
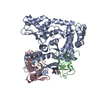
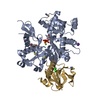
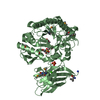
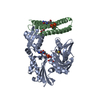
温度: 293 K / 手法: 蒸気拡散法, ハンギングドロップ法 / pH: 7.5 詳細: The ITCH and ubiquitin variant ubv.it.02 were mixed at molarity ratio 1:2, and then concentrated to 17mg/ml. The protein sample was mixed with 1mg/mL chymotrypsin at a 1:1000 (W/W) ...詳細: The ITCH and ubiquitin variant ubv.it.02 were mixed at molarity ratio 1:2, and then concentrated to 17mg/ml. The protein sample was mixed with 1mg/mL chymotrypsin at a 1:1000 (W/W) chymotrypsin:protein ratio right before set up crystallization. Crystal was initially obtained from SGC-I screen condition A05. Crystal used for structure refinement was grown in 1.6M NH4SO4, 0.2M NaAc, 0.1M HEPES pH 7.5, 5% Ethylene Glycol in hanging drop setup, using 1.2uL protein, 1.2uL well solution over 0.5 mL reservoir buffer at 20 C. Crystals grow to mountable size for ~1 weeks. Harvested crystal was flash-frozen in liquid nitrogen. A well solution containing 20% glycerol was used as the cryo-protectant
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード LIGASE (リガーゼ) /
LIGASE (リガーゼ) /  機能・相同性情報
機能・相同性情報
 データ登録者
データ登録者 引用
引用 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード










 PDBj
PDBj




















 集合体
集合体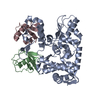



 分子量: 18.015 Da / 分子数: 3 / 由来タイプ: 天然 / 式: H2O
分子量: 18.015 Da / 分子数: 3 / 由来タイプ: 天然 / 式: H2O 試料調製
試料調製 / ビームライン: 19-ID / 波長: 0.97899 Å
/ ビームライン: 19-ID / 波長: 0.97899 Å 解析
解析