 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
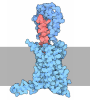
Yorodumi- PDB-4zxb: Structure of the human insulin receptor ectodomain, IRDeltabeta c... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 4zxb | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
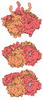
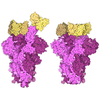
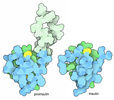
| Title | Structure of the human insulin receptor ectodomain, IRDeltabeta construct, in complex with four Fab molecules | ||||||||||||||||||
 Components Components |
| ||||||||||||||||||
 Keywords Keywords | HORMONE RECEPTOR/IMMUNE SYSTEM / receptor tyrosine kinase extracellular domain antibody fragments / HORMONE RECEPTOR-IMMUNE SYSTEM complex | ||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationregulation of female gonad development / positive regulation of B cell activation / positive regulation of meiotic cell cycle / positive regulation of developmental growth /  insulin-like growth factor II binding / humoral immune response mediated by circulating immunoglobulin / early endosome to late endosome transport / phagocytosis, recognition / male sex determination / exocrine pancreas development ...regulation of female gonad development / positive regulation of B cell activation / positive regulation of meiotic cell cycle / positive regulation of developmental growth / insulin-like growth factor II binding / humoral immune response mediated by circulating immunoglobulin / early endosome to late endosome transport / phagocytosis, recognition / male sex determination / exocrine pancreas development / insulin receptor complex / positive regulation of type IIa hypersensitivity / insulin-like growth factor I binding / insulin receptor activity / regulation of proteolysis / positive regulation of type I hypersensitivity / positive regulation of protein-containing complex disassembly / cargo receptor activity / antibody-dependent cellular cytotoxicity / Fc-gamma receptor I complex binding / dendritic spine maintenance / insulin binding / PTB domain binding / phagocytosis, engulfment / adrenal gland development / IgG immunoglobulin complex / neuronal cell body membrane / Signaling by Insulin receptor / endosome to lysosome transport / IRS activation / activation of protein kinase activity / amyloid-beta clearance / positive regulation of respiratory burst / positive regulation of endocytosis / regulation of embryonic development / positive regulation of receptor internalization / transport across blood-brain barrier / antigen processing and presentation / insulin receptor substrate binding / immunoglobulin mediated immune response / positive regulation of glycogen biosynthetic process / epidermis development / immunoglobulin complex, circulating / Signal attenuation / immunoglobulin receptor binding / phosphatidylinositol 3-kinase binding / positive regulation of phagocytosis / heart morphogenesis / dendrite membrane / Insulin receptor recycling / insulin-like growth factor receptor binding / neuron projection maintenance / positive regulation of glycolytic process / activation of protein kinase B activity / multivesicular body / positive regulation of mitotic nuclear division / receptor-mediated endocytosis / Insulin receptor signalling cascade / B cell differentiation / complement activation, classical pathway / learning / antigen binding / caveola / positive regulation of glucose import / response to bacterium / positive regulation of MAP kinase activity / receptor internalization / receptor protein-tyrosine kinase / memory / cellular response to growth factor stimulus / peptidyl-tyrosine phosphorylation / cellular response to insulin stimulus / positive regulation of nitric oxide biosynthetic process / male gonad development / positive regulation of immune response / late endosome / insulin receptor signaling pathway / glucose homeostasis / amyloid-beta binding / PI5P, PP2A and IER3 Regulate PI3K/AKT Signaling / antibacterial humoral response / protein tyrosine kinase activity / positive regulation of MAPK cascade / protein autophosphorylation / positive regulation of phosphatidylinositol 3-kinase/protein kinase B signal transduction / lysosome / receptor complex / endosome membrane / positive regulation of cell migration / symbiont entry into host cell / positive regulation of protein phosphorylation / G protein-coupled receptor signaling pathway / protein domain specific binding / external side of plasma membrane / axon / protein phosphorylation / positive regulation of cell population proliferation / protein-containing complex binding / regulation of DNA-templated transcription / GTP binding insulin-like growth factor II binding / humoral immune response mediated by circulating immunoglobulin / early endosome to late endosome transport / phagocytosis, recognition / male sex determination / exocrine pancreas development ...regulation of female gonad development / positive regulation of B cell activation / positive regulation of meiotic cell cycle / positive regulation of developmental growth / insulin-like growth factor II binding / humoral immune response mediated by circulating immunoglobulin / early endosome to late endosome transport / phagocytosis, recognition / male sex determination / exocrine pancreas development / insulin receptor complex / positive regulation of type IIa hypersensitivity / insulin-like growth factor I binding / insulin receptor activity / regulation of proteolysis / positive regulation of type I hypersensitivity / positive regulation of protein-containing complex disassembly / cargo receptor activity / antibody-dependent cellular cytotoxicity / Fc-gamma receptor I complex binding / dendritic spine maintenance / insulin binding / PTB domain binding / phagocytosis, engulfment / adrenal gland development / IgG immunoglobulin complex / neuronal cell body membrane / Signaling by Insulin receptor / endosome to lysosome transport / IRS activation / activation of protein kinase activity / amyloid-beta clearance / positive regulation of respiratory burst / positive regulation of endocytosis / regulation of embryonic development / positive regulation of receptor internalization / transport across blood-brain barrier / antigen processing and presentation / insulin receptor substrate binding / immunoglobulin mediated immune response / positive regulation of glycogen biosynthetic process / epidermis development / immunoglobulin complex, circulating / Signal attenuation / immunoglobulin receptor binding / phosphatidylinositol 3-kinase binding / positive regulation of phagocytosis / heart morphogenesis / dendrite membrane / Insulin receptor recycling / insulin-like growth factor receptor binding / neuron projection maintenance / positive regulation of glycolytic process / activation of protein kinase B activity / multivesicular body / positive regulation of mitotic nuclear division / receptor-mediated endocytosis / Insulin receptor signalling cascade / B cell differentiation / complement activation, classical pathway / learning / antigen binding / caveola / positive regulation of glucose import / response to bacterium / positive regulation of MAP kinase activity / receptor internalization / receptor protein-tyrosine kinase / memory / cellular response to growth factor stimulus / peptidyl-tyrosine phosphorylation / cellular response to insulin stimulus / positive regulation of nitric oxide biosynthetic process / male gonad development / positive regulation of immune response / late endosome / insulin receptor signaling pathway / glucose homeostasis / amyloid-beta binding / PI5P, PP2A and IER3 Regulate PI3K/AKT Signaling / antibacterial humoral response / protein tyrosine kinase activity / positive regulation of MAPK cascade / protein autophosphorylation / positive regulation of phosphatidylinositol 3-kinase/protein kinase B signal transduction / lysosome / receptor complex / endosome membrane / positive regulation of cell migration / symbiont entry into host cell / positive regulation of protein phosphorylation / G protein-coupled receptor signaling pathway / protein domain specific binding / external side of plasma membrane / axon / protein phosphorylation / positive regulation of cell population proliferation / protein-containing complex binding / regulation of DNA-templated transcription / GTP bindingSimilarity search - Function | ||||||||||||||||||
| Biological species |  Mus musculus (house mouse) Mus musculus (house mouse) Homo sapiens (human) Homo sapiens (human) | ||||||||||||||||||
| Method | X-RAY DIFFRACTION / SYNCHROTRON / Resolution: 3.3 Å | ||||||||||||||||||
 Authors Authors | Croll, T. / Smith, B.J. / Margetts, M.B. / Whittaker, J. / Weiss, M.A. / Ward, C.W. / Lawrence, M.C. | ||||||||||||||||||
| Funding support |  Australia, 5items Australia, 5items
| ||||||||||||||||||
 Citation Citation | Journal: Structure / Year: 2016 Title: Higher-Resolution Structure of the Human Insulin Receptor Ectodomain: Multi-Modal Inclusion of the Insert Domain. Authors: Croll, T.I. / Smith, B.J. / Margetts, M.B. / Whittaker, J. / Weiss, M.A. / Ward, C.W. / Lawrence, M.C. | ||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 4zxb.cif.gz | 671.8 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb4zxb.ent.gz | 566.6 KB | Display | PDB format |
| PDBx/mmJSON format | 4zxb.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/zx/4zxbftp://data.pdbj.org/pub/pdb/validation_reports/zx/4zxb | HTTPS FTP |
|---|
-Related structure data
| Similar structure data |
|---|






-Links
 PDBj
PDBj


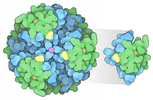






- Assembly
Assembly
| Deposited unit | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||
| Unit cell |
| ||||||||
| Symmetry | Point symmetry: (Schoenflies symbol: C2 (2 fold cyclic)) |
-Components
-Protein , 1 types, 1 molecules E
| #5: Protein | Mass: 102279.945 Da / Num. of mol.: 1 Mutation: Y144H, V731A, T732G, V733N, A734N, residues 735-753 deleted Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Plasmid: pEE14 / Cell line (production host): Lec8 / Production host:  Cricetulus griseus (Chinese hamster) / References: UniProt: P06213 Cricetulus griseus (Chinese hamster) / References: UniProt: P06213 |
|---|
-Antibody , 4 types, 4 molecules ABCD
| #1: Antibody | Mass: 23414.266 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Details: Original Mab obtained from a mouse hybridoma cell line Source: (gene. exp.) Mus musculus (house mouse) / Production host: Mus musculus (house mouse) / References: UniProt: U5LP42 |
|---|---|
| #2: Antibody | Mass: 24385.072 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Details: The original Mab was obtained from a mouse hybridoma Source: (gene. exp.) Mus musculus (house mouse) / Production host: Mus musculus (house mouse) / References: UniProt: P01837 |
| #3: Antibody | Mass: 23622.672 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Details: The original mAb was obtained from a mouse hybridoma Source: (gene. exp.) Mus musculus (house mouse) / Gene: Igh-1a / Production host: Mus musculus (house mouse) / References: UniProt: P01865 |
| #4: Antibody | Mass: 23514.807 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Details: The original mAb was obtained form a mouse hybridoma Source: (gene. exp.) Mus musculus (house mouse) / Gene: Igkc, Igk-C / Production host: Mus musculus (house mouse) / References: UniProt: A2P1G9 |
-Sugars , 5 types, 10 molecules 
| #6: Polysaccharide | / Mass: 570.542 Da / Num. of mol.: 3 Source method: isolated from a genetically manipulated source #7: Polysaccharide | beta-D-mannopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose-(1-4)-2-acetamido-2-deoxy-beta- ...beta-D-mannopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose | / Mass: 586.542 Da / Num. of mol.: 1Source method: isolated from a genetically manipulated source #8: Polysaccharide | 2-acetamido-2-deoxy-beta-D-glucopyranose-(1-2)-alpha-D-mannopyranose-(1-6)-beta-D-mannopyranose-(1- ...2-acetamido-2-deoxy-beta-D-glucopyranose-(1-2)-alpha-D-mannopyranose-(1-6)-beta-D-mannopyranose-(1-4)-2-acetamido-2-deoxy-beta-D-glucopyranose-(1-4)-[alpha-L-fucopyranose-(1-6)]2-acetamido-2-deoxy-beta-D-glucopyranose | / Mass: 1098.016 Da / Num. of mol.: 1Source method: isolated from a genetically manipulated source #9: Polysaccharide | / Mass: 367.349 Da / Num. of mol.: 2Source method: isolated from a genetically manipulated source #10: Sugar | N-Acetylglucosamine Type: D-saccharide, beta linking / Mass: 221.208 Da / Num. of mol.: 3 Type: D-saccharide, beta linking / Mass: 221.208 Da / Num. of mol.: 3Source method: isolated from a genetically manipulated source Formula: C8H15NO6 |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 4.95 Å3/Da / Density % sol: 75.15 % |
|---|---|
| Crystal grow | Temperature: 293 K / Method: vapor diffusion, hanging drop / pH: 6.5 Details: Crystals were produced by vapour diffusion using 1 microlitre of the complex (2.6 mg/ml) in 10 mM HEPES (pH 7.5), 0.02% sodium azide and 10% D-trehalose solution and 1 microlitres of well ...Details: Crystals were produced by vapour diffusion using 1 microlitre of the complex (2.6 mg/ml) in 10 mM HEPES (pH 7.5), 0.02% sodium azide and 10% D-trehalose solution and 1 microlitres of well solution containing 10% PEG 8000, 0.1 M MES (pH 6.5) and 0.1 M MgAc2. |
-Data collection
| Diffraction | Mean temperature: 100 K |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: Photon Factory  / Beamline: BL-5A / Wavelength: 1 Å / Beamline: BL-5A / Wavelength: 1 Å |
| Detector | Type: ADSC QUANTUM 315 / Detector: CCD / Date: Jun 22, 2006 |
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 1 Å / Relative weight: 1 |
| Reflection | Resolution: 3.3→35 Å / Num. obs: 57953 / % possible obs: 97.9 % / Redundancy: 7 % / Biso Wilson estimate: 90.48 Å2 / Net I/σ(I): 11.39 |
| Reflection shell | Resolution: 3.3→3.4 Å / Redundancy: 3.6 % / Mean I/σ(I) obs: 0.78 / % possible all: 86.6 |
- Processing
Processing
| Software |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Resolution: 3.3→34.46 Å / Cor.coef. Fo:Fc: 0.9258 / Cor.coef. Fo:Fc free: 0.9127 / SU R Cruickshank DPI: 0.978 / Cross valid method: THROUGHOUT / σ(F): 0 / SU R Blow DPI: 0.854 / SU Rfree Blow DPI: 0.359 / SU Rfree Cruickshank DPI: 0.37
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 134.12 Å2
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine analyze | Luzzati coordinate error obs: 0.912 Å | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 3.3→34.46 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell | Resolution: 3.3→3.39 Å / Total num. of bins used: 20
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS params. | Method: refined / Refine-ID: X-RAY DIFFRACTION
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS group |
|
