手法: 溶液NMR 詳細: IPAP EXPERIMENTS FOR DIPOLAR COUPLINGS. DIPOLAR COUPLINGS WERE MEASURED IN A NEMATIC PHASE OF A 4-5% PEG/HEXANOL (SURFACTANT TO ALCOHOL RATION OF 0.96)
NMR実験
Conditions-ID
Experiment-ID
Solution-ID
タイプ
1
1
1
TRIPLERESONANCEFORASSIGNMENTOFPROTEIN
1
2
1
QUANTITATIVEJCORRELATIONFORCOUPLINGCONSTANTS
1
3
1
3D, 4DHETERONUCLEARSEPARATED, FILTEREDNOEEXPTS
1
4
1
IPAP EXPERIMENTS FOR DIPOLAR COUPLINGS
-
試料調製
試料状態
イオン強度: 10 mM SODIUM PHOSPHATE / pH: 7.0 / 温度: 308.00 K
手法: CONJOINED RIGID BODY, TORSION ANGLE DYNAMICS / ソフトェア番号: 1 詳細: THE STRUCTURES WERE CALCULATED BY CONJOINED RIGID BODY/TORSION ANGLE DYNAMICS (SCHWIETERS & CLORE (2001) J.MAGN.RESON 152, 288-302; (CLORE & BEWLEY (2002) J.MAGN.RESON. 154, 329-335) THE ...詳細: THE STRUCTURES WERE CALCULATED BY CONJOINED RIGID BODY/TORSION ANGLE DYNAMICS (SCHWIETERS & CLORE (2001) J.MAGN.RESON 152, 288-302; (CLORE & BEWLEY (2002) J.MAGN.RESON. 154, 329-335) THE TARGET FUNCTIONS COMPRISES TERMS FOR THE NOE RESTRAINTS, THE SIDECHAIN TORSION ANGLE RESTRAINTS, THE BACKBONE TORSION ANGLE RESTRAINTS FOR 4 VARIABLE REGIONS OF IIAMTL, THE DIPOLAR COUPLING RESTRAINTS (CLORE ET AL. J.MAGN.RESON. 131, 159-162 (1998); J.MAGN.RESON. 133, 216-221(1998)), THE RADIUS OF GYRATION (KUSZEWSKI ET AL. (1999), AND A QUARTIC VAN DER WAALS REPULSION TERM (NILGES ET AL. (1988) FEBS LETT. 229, 129- 136). THE STARTING COORDINATES COME FROM THE X-RAY STRUCTURES (WITH PROTONS ADDED) OF E. COLI HPR (1POH, JIA ET AL. (1993) J.BIOL.CHEM. 268, 22940-22501, RESOLUTION 1.5 A); AND IIAMTL (MOLECULE D OF 1A3A, VAN MONTFORT ET AL. STRUCTURE 5, 217-225 (1998); RESOLUTION 1.8A). SEVERAL DIFFERENT INITIAL ORIENTATIONS OF THE TWO PROTEINS WERE EMPLOYED WITH THE CA-CA DISTANCE BETWEEN THE ACTIVE SITE HISTIDINES RANGING FROM 28 TO 95 A, INCLUDING ORIENTATIONS WHERE THE TWO ACTIVE SITE HISTIDINES ARE NOT OPPOSED AND WHERE HPR IS DIRECTED TOWARDS THE FACE OF IIAMTL OPPOSITE TO THE IIAMTL ACTIVE SITE. THE BACKBONE COORDINATES AND NON-INTERFACIAL SIDECHAINS (EXCLUDING THE FOUR VARIABLE REGIONS OF IIAMTL: RESIDUES 51-54, 66-78, 91-96 AND 104-110) ARE TREATED AS RIGID BODIES THROUGHOUT WITH IIAMTL HELD FIXED, HPR ALLOWED TO ROTATE AND TRANSLATE, AND THE AXIS OF THE DIPOLAR COUPLING ALIGNMENT TENSOR FREE TO ROTATE. THE INTERFACIAL SIDECHAINS, AS WELL AS THE BACKBONE AND SIDECHAINS OF THE FOUR VARIABLE REGIONS OF IIAMTL, ARE GIVEN FULL TORSIONAL DEGREES OF FREEDOM. ALSO NOTE THAT GLU59 AND HIS111 ARE REFINED IN TWO ALTERNATE CONFORMATIONS. IN THIS ENTRY THE LAST COLUMN REPRESENTS THE AVERAGE RMS DIFFERENCE BETWEEN THE INDIVIDUAL SIMULATED ANNEALING STRUCTURES AND THE MEAN COORDINATE POSITIONS. IT IS IMPORTANT TO NOTE THAT THE VALUES GIVEN FOR THE BACKBONE ATOMS AND NON-INTERFACIAL SIDECHAINS (EXCLUDING THE FOUR VARIABLE REGIONS OF IIAMTL) PROVIDE ONLY A MEASURE OF THE PRECISION WITH WHICH THE RELATIVE ORIENTATION OF THE TWO PROTEINS HAVE BEEN DETERMINED AND DOES NOT TAKE INTO ACCOUNT THE ERRORS IN THE X-RAY COORDINATES OF HPR AND IIAMTL. RESIDUE NUMBERING: IIAMTL: 4-147 (RESIDUES 1-3 ARE DISORDERED IN SOLUTION AND NOT VISIBLE IN THE ELECTRON DENSITY MAP OF THE CRYSTAL STRUCTURE OF THE FREE PROTEIN). HPR: 301-385 (CORRESPONDING TO RESIDUES 1-85). PHOSPHATE: RESIDUE 200 THREE SETS OF COORDINATES ARE GIVEN: MODEL 1: RESTRAINED REGULARIZED MEAN COORDINATES OF THE UNPHOSPHORYLATED HPR-IIAGLC COMPLEX SOLVED ON THE BASIS OF 107 INTERMOLECULAR INTERPROTON DISTANCE DISTANCE RESTRAINTS, 105 INTRAMOLECULAR DISTANCE RESTRAINTS (RELATING TO INTERFACIAL SIDECHAINS, AS WELL AS THE FOUR VARIABLE REGIONS OF IIAMTL), 70 INTERFACIAL SIDECHAIN TORSION ANGLE RESTRAINTS, 62 TORSION ANGLE RESTRAINTS FOR THE VARIABLE REGIONS OF IIAMTL, AND 528 RESIDUAL DIPOLAR COUPLINGS. CROSS-VALIDATION WAS USED FOR THE DIPOLAR COUPLINGS (CLORE AND GARRETT (1999) J. AM. CHEM. SOC. 121, 9008-9012). MODEL 2: RESTRAINED REGULARIZED MEAN COORDINATES FOR THE MODEL OF THE DISSOCIATIVE PHOSPHORYL TRANSITION STATE HPR-IIAMTL COMPLEX. EXPERIMENTAL RESTRAINTS ARE IDENTICAL TO THOSE USED FOR MODEL 3, BUT COVALENT GEOMETRY RESTRAINTS ARE INCLUDED RELATING TO THE PENTACOORDINATE PHOSPHORYL GROUP IN A TRIGONAL BIPYRAMIDAL GEOMETRY. THE STRUCTURE IS DERIVED FROM MODEL 3 BY RESTRAINED MINIMIZATION. THE N-P BOND LENGTHS ARE RESTRAINED TO 3 A. THE CA-CA DISTANCE BETWEEN HIS315 (HPR) AND HIS65 (IIAMTL) REMAINS ESSENTIALLY UNCHANGED FROM MODEL 3, BUT THE ND1-NE2 DISTANCE BETWEEN HIS315 AND HIS65 IS REDUCED TO 6 A, WITH ESSENTIALLY IDEALIZED GEOMETRY OF THE PHOSPHORYL TRANSITION STATE. THE ND1-NE2 DISTANCE CORRESPONDS TO A DISSOCIATIVE TRANSITION STATE. THE RMS DIFFERENCE BETWEEN THE MEAN STRUCTURE OF THE UNPHOSPHORYLATED COMPLEX (MODEL 3) AND THE TRANSITION STATE COMPLEX IS 0.2 A FOR BACKBONE COORDINATES IMMEDIATELY ADJACENT TO THE ACTIVE SITE HISTIDINES (RESIDUES 64-66 AND RESIDUES 316-317). THE REMAINING BACKBONE COORDINATES DO NOT SHIFT. MODEL 3: RESTRAINED REGULARIZED MEAN COORDINATES FOR THE MODEL OF THE ASSOCIATIVE PHOSPHORYL TRANSITION STATE HPR-IIAGLC COMPLEX. CALCULATED LIKE MODEL 2 BUT WITH THE N-P BOND LENGTHS RESTRAINED TO 2A. THE STRUCTURE IS DERIVED FROM MODEL 1 BY RESTRAINED MINIMIZATION. THE RMS DIFFERENCE BETWEEN THE MEAN STRUCTURES OF THE UNPHOSPHORYLATED COMPLEX (MODEL 1) AND THE TRANSITION STATE COMPLEX IS 0.4 A FOR BACKBONE COORDINATES IMMEDIATELY ADJACENT TO THE ACTIVE SITE HISTIDINES (RESIDUES 64-66 AND RESIDUES 316-317). THE REMAINING BACKBONE COORDINATES DO NOT SHIFT. HPR-IIAMTL COMPLEX DEVIATIONS FROM IDEALIZED GEOMETRY: BONDS 0.006 A, ANGLES 0.82 DEG, IMPROPER TORSIONS 0.97 DEG RMS DEVIATIONS FROM NOE DISTANCE RESTRAINTS: 0.007 A RMS DEVIATIONS FROM SIDECHAIN TORSION ANGLE RESTRAINTS: 0.26 DEG. RMS DEVIATIONS FROM BACKBONE TORSION ANGLE RESTRAINTS: 1.2 DEG. DIPOLAR COUPLING R-FACTORS (CLORE AND GARRETT (1999) J. AM. CHEM. SOC. 121, 9008-9012): HPR IIAMTL NH 19.1% 19.2% CaH 25.9% 18.7% NC' 34.0% 32.1% [NOTE ONE ALIGNMENT TENSOR IS USED FOR THE NH DIPOLAR COUPLINGS (FOR BOTH HPR AND IIAMTL), AND ANOTHER FOR THE CAH AND NC' DIPOLAR COUPLINGS (FOR BOTH HPR AND IIAMTL), SINCE THE LATTER SET OF DIPOLAR COUPLINGS WERE OBTAINED FROM A DIFFERENT BATCH OF PEG/HEXANOL THAN THE FORMER. THE ORIENTATION OF THE TWO ALIGNMENT TENSORS DIFFERS BY ONLY 1.9 DEG. NOTE THE ALIGNMENT TENSORS FOR HPR AND IIAMTL ARE THE SAME. FOR REFERENCE THE DIPOLAR COUPLING R-FACTORS FOR THE FREE STRUCTURES (USING INDIVIDUAL ALIGNMENT TENSORS FOR THE TWO PROTEINS) ARE 21.3% (NH), 21.1% (CaH), 33.6% (NC') FOR THE X-RAY STRUCTURE OF HPR, AND 19.2% (NH), 18.0% (CaH) AND 32.0% (NC') FOR THE RESTRAINED REGULARIZED MEAN STRUCTURE OF IIAMTL IN THE COMPLEX].
NMRアンサンブル
コンフォーマー選択の基準: REGULARIZED MEAN STRUCTURES 計算したコンフォーマーの数: 200 / 登録したコンフォーマーの数: 3
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード TRANSFERASE (転移酵素) /
TRANSFERASE (転移酵素) /  機能・相同性情報
機能・相同性情報
 データ登録者
データ登録者 引用
引用 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード



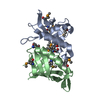
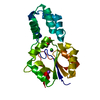
 PDBj
PDBj 集合体
集合体
 分子量: 78.972 Da / 分子数: 1 / 由来タイプ: 合成 / 式: PO3
分子量: 78.972 Da / 分子数: 1 / 由来タイプ: 合成 / 式: PO3 試料調製
試料調製 解析
解析