ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: EMDB / ID: EMD-10120 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
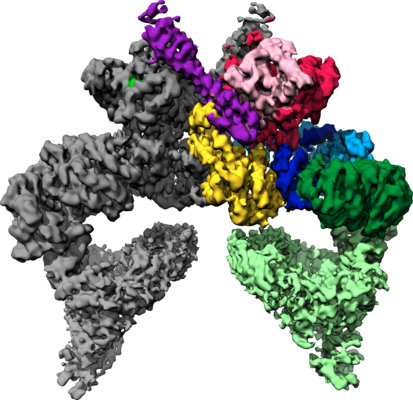
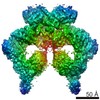
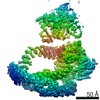
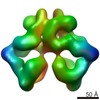
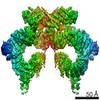
| タイトル | Structure of nucleotide-bound Tel1/ATM | |||||||||
 マップデータ マップデータ | Tel1 Whole Dimer | |||||||||
 試料 試料 |
| |||||||||
 キーワード キーワード |  Kinase (キナーゼ) / DNA Damage Response / CryoEM (低温電子顕微鏡法) / Phosphatidylinositol-3-kinase-like kinase / HYDROLASE (加水分解酵素) Kinase (キナーゼ) / DNA Damage Response / CryoEM (低温電子顕微鏡法) / Phosphatidylinositol-3-kinase-like kinase / HYDROLASE (加水分解酵素) | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報DNA Damage/Telomere Stress Induced Senescence / Sensing of DNA Double Strand Breaks / Pexophagy / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / telomeric DNA binding / signal transduction in response to DNA damage / negative regulation of TORC1 signaling / telomere maintenance / DNA damage checkpoint signaling / double-strand break repair ...DNA Damage/Telomere Stress Induced Senescence / Sensing of DNA Double Strand Breaks / Pexophagy / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / telomeric DNA binding / signal transduction in response to DNA damage / negative regulation of TORC1 signaling / telomere maintenance / DNA damage checkpoint signaling / double-strand break repair / chromatin organization / chromosome, telomeric region / non-specific serine/threonine protein kinase / protein kinase activity / リン酸化 / protein serine kinase activity / DNA修復 / protein serine/threonine kinase activity / DNA damage response / ミトコンドリア / ATP binding / 細胞核類似検索 - 分子機能 | |||||||||
| 生物種 |  Saccharomyces cerevisiae (パン酵母) Saccharomyces cerevisiae (パン酵母) | |||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 4.0 Å | |||||||||
 データ登録者 データ登録者 | Yates LA / Williams RM | |||||||||
| 資金援助 |  英国, 1件 英国, 1件
| |||||||||
 引用 引用 | ジャーナル: Structure / 年: 2020 タイトル: Cryo-EM Structure of Nucleotide-Bound Tel1 Unravels the Molecular Basis of Inhibition and Structural Rationale for Disease-Associated Mutations. 著者: Luke A Yates / Rhys M Williams / Sarem Hailemariam / Rafael Ayala / Peter Burgers / Xiaodong Zhang /  要旨: Yeast Tel1 and its highly conserved human ortholog ataxia-telangiectasia mutated (ATM) are large protein kinases central to the maintenance of genome integrity. Mutations in ATM are found in ataxia- ...Yeast Tel1 and its highly conserved human ortholog ataxia-telangiectasia mutated (ATM) are large protein kinases central to the maintenance of genome integrity. Mutations in ATM are found in ataxia-telangiectasia (A-T) patients and ATM is one of the most frequently mutated genes in many cancers. Using cryoelectron microscopy, we present the structure of Tel1 in a nucleotide-bound state. Our structure reveals molecular details of key residues surrounding the nucleotide binding site and provides a structural and molecular basis for its intrinsically low basal activity. We show that the catalytic residues are in a productive conformation for catalysis, but the phosphatidylinositol 3-kinase-related kinase (PIKK) regulatory domain insert restricts peptide substrate access and the N-lobe is in an open conformation, thus explaining the requirement for Tel1 activation. Structural comparisons with other PIKKs suggest a conserved and common allosteric activation mechanism. Our work also provides a structural rationale for many mutations found in A-T and cancer. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | EMマップ: SurfViewMolmilJmol/JSmol |
| 添付画像 |
- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_10120.map.gz | 190.1 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-10120-v30.xmlemd-10120.xml | 21.3 KB 21.3 KB | 表示 表示 | EMDBヘッダ |
| FSC (解像度算出) | emd_10120_fsc_1.xmlemd_10120_fsc_2.xmlemd_10120_fsc_3.xml | 13.5 KB 13.5 KB 13.5 KB | 表示 表示 表示 | FSCデータファイル |
| 画像 | 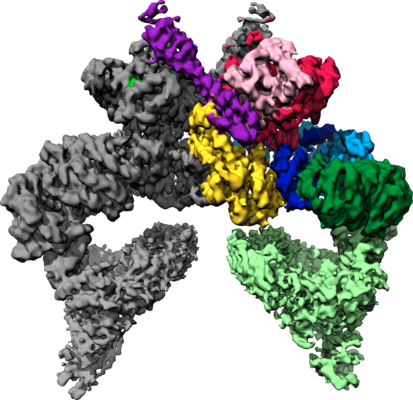 emd_10120.png emd_10120.png | 210.5 KB | ||
| Filedesc metadata | emd-10120.cif.gz | 6.5 KB | ||
| その他 | emd_10120_additional_1.map.gzemd_10120_additional_2.map.gz | 193.6 MB 193.6 MB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-10120ftp://ftp.pdbj.org/pub/emdb/structures/EMD-10120 http://ftp.pdbj.org/pub/emdb/structures/EMD-10120ftp://ftp.pdbj.org/pub/emdb/structures/EMD-10120 | HTTPS FTP |
-関連構造データ




-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| 「今月の分子」の関連する項目 |


-マップ
| ファイル | ダウンロード / ファイル: emd_10120.map.gz / 形式: CCP4 / 大きさ: 209.3 MB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | Tel1 Whole Dimer | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ボクセルのサイズ | X=Y=Z: 1.085 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 詳細 | EMDB XML:
CCP4マップ ヘッダ情報:
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
-添付データ
-追加マップ: Tel1 Kinase Core Dimer
| ファイル | emd_10120_additional_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | Tel1 Kinase Core Dimer | ||||||||||||
| 投影像・断面図 |
| ||||||||||||
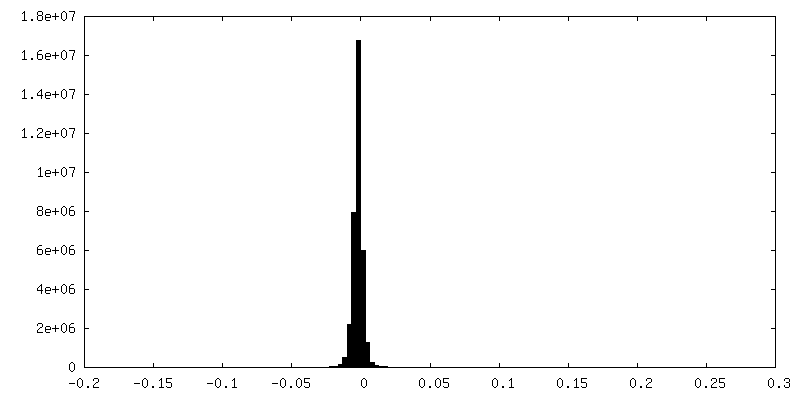
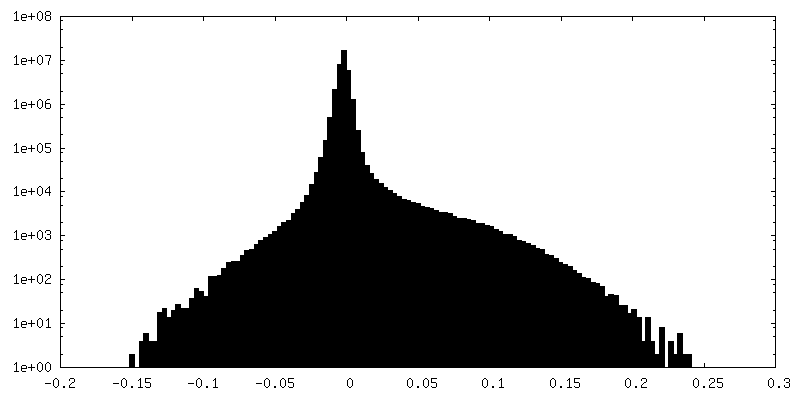
| 密度ヒストグラム |
 Z
Z Y
Y X
X







-追加マップ: Tel1 FATKIN Dimer
| ファイル | emd_10120_additional_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | Tel1 FATKIN Dimer | ||||||||||||
| 投影像・断面図 |
| ||||||||||||
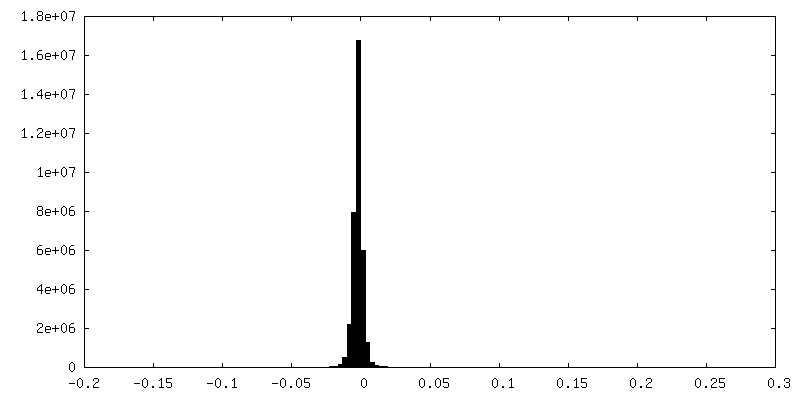
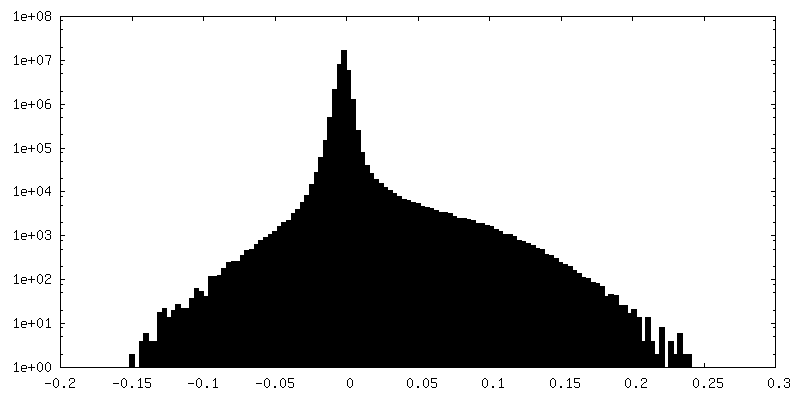
| 密度ヒストグラム |






- 試料の構成要素
試料の構成要素
-全体 : Tel1/ATM
| 全体 | 名称: Tel1/ATM |
|---|---|
| 要素 |
|
-超分子 #1: Tel1/ATM
| 超分子 | 名称: Tel1/ATM / タイプ: complex / ID: 1 / 親要素: 0 / 含まれる分子: #1 |
|---|---|
| 由来(天然) | 生物種: Saccharomyces cerevisiae (パン酵母) |
-分子 #1: Serine/threonine-protein kinase TEL1,Serine/threonine-protein kin...
| 分子 | 名称: Serine/threonine-protein kinase TEL1,Serine/threonine-protein kinase TEL1,Serine/threonine-protein kinase TEL1,Serine/threonine-protein kinase TEL1,Serine/threonine-protein kinase TEL1 タイプ: protein_or_peptide / ID: 1 / コピー数: 2 / 光学異性体: LEVO / EC番号: non-specific serine/threonine protein kinase |
|---|---|
| 由来(天然) | 生物種: Saccharomyces cerevisiae (パン酵母) |
| 分子量 | 理論値: 292.579406 KDa |
| 組換発現 | 生物種: Saccharomyces cerevisiae (パン酵母) |
| 配列 | 文字列: (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) ...文字列: (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)GS IRGGKQRVFA TFIKCLQKLD SSNIINIMNS ISSYMAQV S YKNQSIIFYE IKSLFGPPQQ SIEKSAFYSL AMSMLSLVSY PSLVFSLEDM MTYSGFNHTR AFIQQALNKI TVAFRYQNL TELFEYCKFD LIMYWFNRTK VPTSKLEKEW DISLFGFADI HEFLGRYFVE ISAIYFSQGF NQKWILDMLH AITGNGDAYL VDNSYYLCI PLAFISGGVN ELIFDILPQI SGKTTVKYHK KYRLLMLKWI IRFTDLGSLT ELRSTVEKLF PTSYLSPYLF E NSSVSMRY QYPLHIPLAL GATLVQTQFA HEKNNTHEFK LLFLSVITDL EKTSTYIGKL RCARELKYLF VLYENVLVKS ST LNFIIIR LSKFLIDTQI HDEVITIFSS LLNLADKNTF EIEPSLPNLF CKIFIYLREN KQLSPSFQQA IKLLEHRDLI KIK TWKYCL DAIFGNIVQD DIYENTELLD ASDCGVDDVV LVSLLFSYAR RPVASKIGCS LSKAAAINIL KHHVPKEYLS KNFK LWFAA LSRRILQQEV QRERSTNFNN EVHLKNFEMV FRHPEQPHMI YQRISTFNKE AELYDSTEVF FISECILTYL VGYSI GNSE SEFCFRDNIM NENKDKVAPL DKDVLNAIYP LANNFGMESF ICDTYLSVNE PYNCWLSKFA RSLIHQISFN IPPIVC LYP LCKGSTAFCE LVLTDLFFLS TTYDPKSCLN WSNRIFTQIA MLLHVKDSEI KLKMLFNVIK MIRMGSRCKE RNCLRIY SS LDLQEICQIS LKIKEFKFGY LLFEEMNMPN IREMNINTLQ KIYECINDGD FLAGLPVPHS IEGVLNSINR IDSDTWKR F LFNNADFDAN YTTSLEEEKE SLIKATEDSG FYGLTSLLES RLSGSSDVYK WNLELGDWKL LTPKVVDSKA KGLYYAIKN LPQDVGFAEK SLEKSLLTIF DSRQHFISQT EWMDTLNAII EFIKIAAIPQ DVTSFPQTLM SIMKADKERL NTIDFYDHKT TLKSRHTLM NVLSRNSLDE NVKCSKYLRL GSIIQLANYV QLAIANGAPQ DALRNATLMS KTVKNIAKLY DDPSVVSQIE K LASFTSAN ALWESREYKA PVMIMRDLLA QNEKNISESI LYDDFKLLIN VPMDQIKARL VKWSSESRLE PAAAIYEKII VN WDINVED HESCSDVFYT LGSFLDEQAQ KLRSNGEIED REHRSYTGKS TLKALELIYK NTKLPENERK DAKRHYNRVL LQY NRDSEV LKALLLQKEK FLWHALHFYL NTLVFSNRYD NDIIDKFCGL WFENDDNSKI NQLLYKEIGT IPSWKFLPWV NQIA SKISM EENEFQKPLQ LTMKRLLYKL PYDSLYSVMS ILLYEKQSNK DTNISQKIQA VKKILLELQG YDRGAFAKKY LLPVQ EFCE MSVELANLKF VQNTKTLRLA NLKIGQYWLK QLNMEKLPLP TSNFTVKSSA DGRKARPYIV SVNETVGITT TGLSLP KIV TFNISDGTTQ KALMKGSNDD LRQDAIMEQV FQQVNKVLQN DKVLRNLDLG IRTYKVVPLG PKAGIIEFVA NSTSLHQ IL SKLHTNDKIT FDQARKGMKA VQTKSNEERL KAYLKITNEI KPQLRNFFFD SFPDPLDWFE AKKTYTKGVA ASSIVGYI L GLGDRHLNNI LLDCSTGEPI HIDLGIAFDQ GKLLPIPELV PFRLTRDIVD GFGVTGVDGL FRRSCERVYA VLRKDYVKV MCVLNILKWD PLYSWVMSPV KKYEHLFEEE HEITNFDNVS KFISNNDRNE NQESYRALKG VEEKLMGNGL SVESSVQDLI QQATDPSNL SVIYMGWSPF Y UniProtKB: Serine/threonine-protein kinase TEL1 |
-分子 #2: PHOSPHOAMINOPHOSPHONIC ACID-ADENYLATE ESTER
| 分子 | 名称: PHOSPHOAMINOPHOSPHONIC ACID-ADENYLATE ESTER / タイプ: ligand / ID: 2 / コピー数: 2 / 式: ANP |
|---|---|
| 分子量 | 理論値: 506.196 Da |
| Chemical component information | 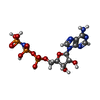 ChemComp-ANP: |
-分子 #3: MAGNESIUM ION
| 分子 | 名称: MAGNESIUM ION / タイプ: ligand / ID: 3 / コピー数: 2 / 式: MG |
|---|---|
| 分子量 | 理論値: 24.305 Da |
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | particle |
-試料調製
| 緩衝液 | pH: 7.4 |
|---|---|
| 凍結 | 凍結剤: ETHANE |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | FEI TITAN KRIOS |
|---|---|
| 電子線 | 加速電圧: 300 kV / 電子線源: FIELD EMISSION GUN |
| 電子光学系 | 照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELDBright-field microscopy |
| 撮影 | フィルム・検出器のモデル: FEI FALCON III (4k x 4k) 平均電子線量: 88.8 e/Å2 |
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
-画像解析
| 初期モデル | モデルのタイプ: EMDB MAP EMDB ID: |
|---|---|
| 初期 角度割当 | タイプ: MAXIMUM LIKELIHOOD / ソフトウェア - 名称: RELION (ver. 3.0) |
| 最終 角度割当 | タイプ: MAXIMUM LIKELIHOOD / ソフトウェア - 名称: RELION (ver. 3.0) |
| 最終 再構成 | 想定した対称性 - 点群: C2 (2回回転対称) / 解像度のタイプ: BY AUTHOR / 解像度: 4.0 Å / 解像度の算出法: FSC 0.143 CUT-OFF / ソフトウェア - 名称: RELION (ver. 3.0) / 使用した粒子像数: 167596 |
| FSC曲線 (解像度の算出) | 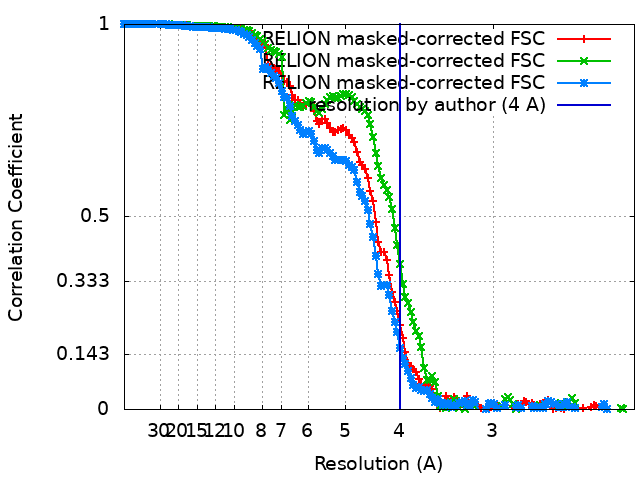 |